Przeczytaj
Fotosynteza typu CIndeks dolny 33
Większość gatunków roślin to rośliny prowadzące fotosyntezę typu CIndeks dolny 33. U roślin tych występuje jednoetapowy mechanizm asymilacjiasymilacji dwutlenku węgla – w cyklu Calvina.
Więcej informacji na temat redukcji węgla w cyklu Calvina znajdziesz tutajtutaj.
Rośliny typu CIndeks dolny 33 zasiedlają tereny o chłodnym i umiarkowanym klimacie. Charakterystycznymi cechami budowy anatomicznej liści u większości roślin typu CIndeks dolny 33 są zróżnicowanie miękiszu asymilacyjnegomiękiszu asymilacyjnego na palisadowy i gąbczasty oraz ruchy aparatów szparkowych, które pozostają otwarte w ciągu dnia, a zamknięte w nocy. Zatem wymiana gazowa, w wyniku której rośliny uzyskują dwutlenek węgla niezbędny do przebiegu fazy ciemnejfazy ciemnej, zachodzi w tym samym czasie co faza jasnafaza jasna fotosyntezy. Pierwotnym akceptorem COIndeks dolny 22 jest związek pięciowęglowy – rybulozo‑1,5‑bisfosforan (RuBP). Reakcję karboksylacjikarboksylacji katalizuje enzym RuBisCO. Pierwszym trwałym produktem karboksylacji jest kwas 3‑fosfoglicerynowy (PGA) – związek trójwęglowy, stąd nazwa grupy roślin i typu fotosyntezy – CIndeks dolny 33.
Szczególne typy fotosyntezy: CIndeks dolny 44 i CAM
Rośliny typu CIndeks dolny 44 i CAM wykształciły inne szlaki fotosyntetyczne niż rośliny CIndeks dolny 33. Różnice metaboliczne są wyrazem przystosowania do wymagających warunków środowiska. U roślin CIndeks dolny 44 i CAM istnieje dwuetapowy mechanizm asymilacji dwutlenku węgla.
Zatem u roślin typu CIndeks dolny 44 i typu CAM:
pierwotnym akceptorem dwutlenku węgla jest fosfoenolopirogronian (PEP), a pierwszym trwałym produktem karboksylacji – szczawiooctan (OAA);
wtórnym akceptorem dwutlenku węgla jest rybulozo‑1,5‑bisfosforan (RuBP).
Fotosynteza typu CIndeks dolny 44
Fotosynteza CIndeks dolny 44 to szczególny szlak fotosyntetyczny występujący u roślin strefy okołorównikowej i zwrotnikowej. Rośliny te są przystosowane do znoszenia wysokich temperatur, dużego natężenia światła i ograniczonej dostępności wody. Fotosynteza typu CIndeks dolny 44 występuje u wielu niespokrewnionych ze sobą gatunków roślin, tworzących jedną grupę – roślin typu CIndeks dolny 44, których wspólną cechą jest przeprowadzanie fotosyntezy typu CIndeks dolny 44.
Przykłady roślin przeprowadzających fotosyntezę typu CIndeks dolny 44 (rośliny typu CIndeks dolny 44)





Rośliny typu CIndeks dolny 44 zajmują siedliska klimatu gorącego, dlatego prowadzą oszczędną gospodarkę wodną. W ciągu dnia przymykają aparaty szparkowe, co pozwala ograniczyć straty wody na drodze transpiracjitranspiracji. Jednocześnie następuje zmniejszenie wydajności wymiany gazowej między wnętrzem rośliny a atmosferą. Zmniejszona dostępność dwutlenku węgla nie ogranicza jednak natężenia procesu fotosyntezy. Dzieje się tak dlatego, że karboksylaza fosfoenolopirogronianowa ma duże powinowactwo do dwutlenku węgla i może go wydajnie wiązać nawet przy niewielkim stężeniu tego gazu.
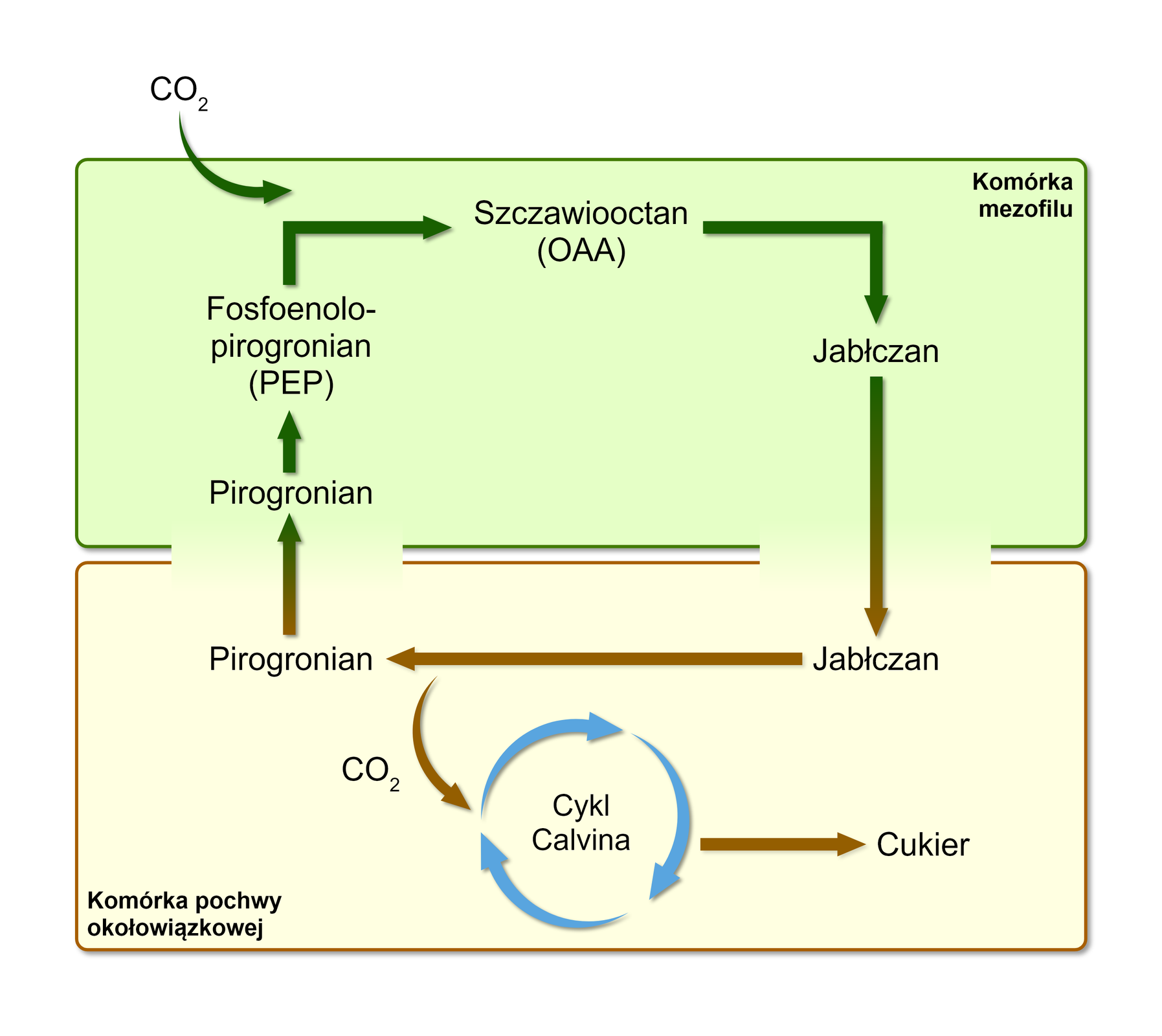
Liście roślin typu CIndeks dolny 44 zawierają niezróżnicowany miękisz asymilacyjny (mezofil), którego komórki są luźno rozmieszczone między górną i dolną skórką liścia. Oprócz komórek mezofilu w liściu obecne są również komórki pochwy okołowiązkowej, które ściśle otaczają wiązkę przewodzącą.

W ciągu dnia w cytoplazmie komórek miękiszu asymilacyjnego (mezofilu) zachodzi przyłączenie dwutlenku węgla do związku trójwęglowego – fosfoenolopirogronianu (PEP), w wyniku czego powstaje związek czterowęglowy – szczawiooctan (OAA). Następnie szczawiooctan jest redukowany do jabłczanu, który przez plazmodesmyplazmodesmy przenika do komórek pochwy okołowiązkowej. Na terenie ich cytoplazmy dochodzi do odłączenia cząsteczki dwutlenku węgla z jabłczanu i powstania trójwęglowego pirogronianiu. Uwolniony dwutlenek węgla zostaje przyłączony do rybulozo‑1,5‑bisfosforanu (RuBP) i podlega przemianom cyklu Calvina, tak samo jak w fotosyntezie typu CIndeks dolny 33. Pirogronian przenika do komórek miękiszu asymilacyjnego i zostaje wykorzystany do odtworzenia fosfoenolopirogronianu.
Fotosynteza typu CAM
Fotosynteza CAM (ang. crassulacean acid metabolism, metabolizm kwasowy gruboszowatych) to szczególny szlak fotosyntetyczny po raz pierwszy odkryty u roślin z rodziny gruboszowatych (Crassulaceae). Fotosynteza CAM występuje także u innych gatunków roślin, np. u wanilii (Vanilla), ananasa jadalnego (Ananas comosus) i większości kaktusów (Cactaceae). Rośliny typu CAM to niespokrewniona ze sobą grupa, której cechą wspólną jest obecność fotosyntezy typu CAM.
Przykłady roślin przeprowadzających fotosyntezę typu CAM (rośliny typu CAM)





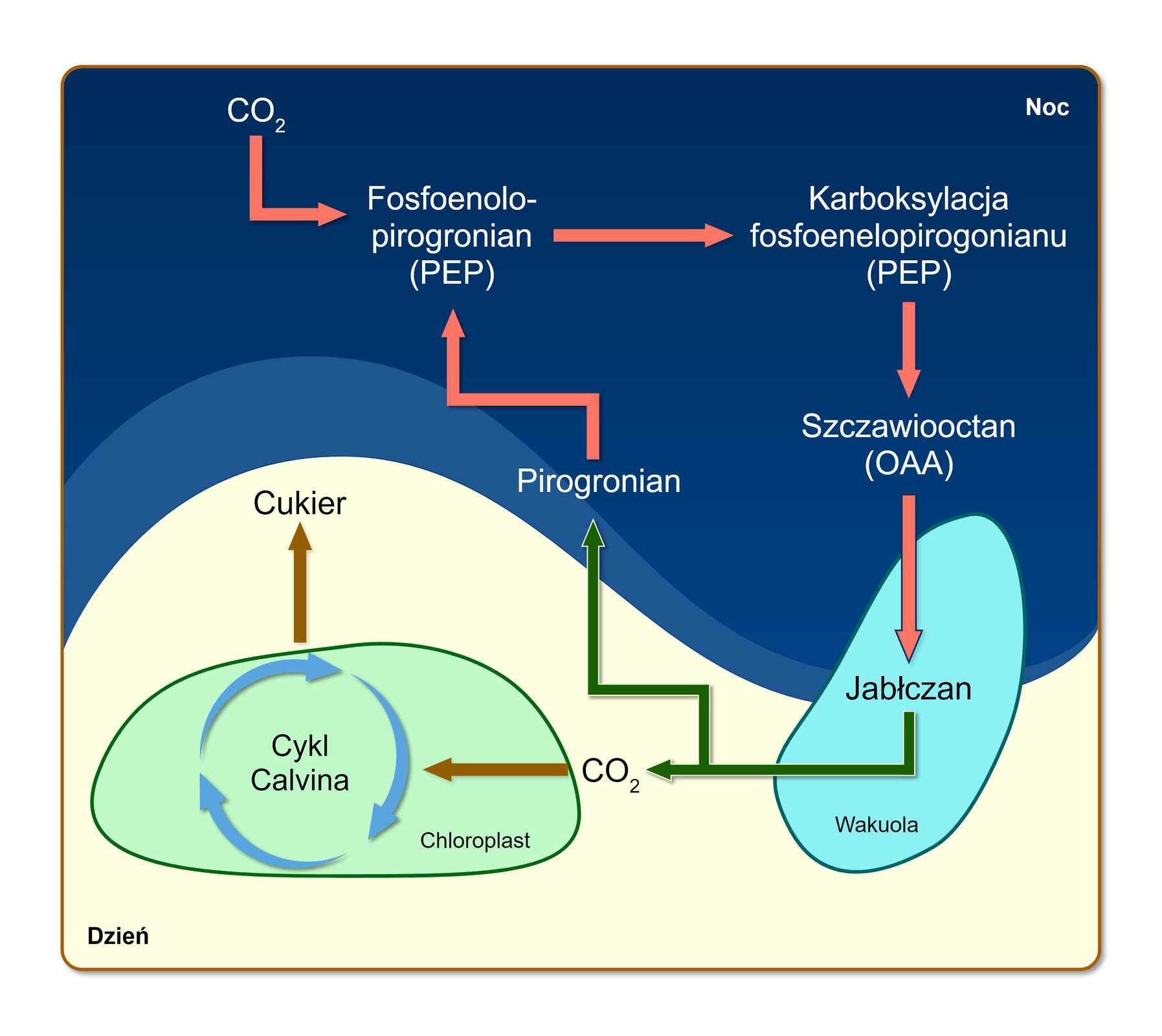
Rośliny typu CAM zajmują siedliska ubogie w wodę, np. wysokogórskie rumowiska skalne, półpustynie lub piaszczyste tereny pustyń. Prowadzą bardzo oszczędną gospodarkę wodną, dlatego w ciągu dnia przy wysokich temperaturach powietrza zamykają aparaty szparkowe. Zamknięcie aparatów szparkowych ogranicza straty wody na drodze transpiracji, jednocześnie uniemożliwiając wymianę gazową między wnętrzem rośliny a atmosferą. Brak możliwości pozyskiwania dwutlenku węgla w dzień sprawia, że rośliny typu CAM asymilują COIndeks dolny 22 w nocy, kiedy aparaty szparkowe pozostają otwarte.
W nocy aparaty szparkowe roślin typu CAM są otwarte. Wówczas rośliny te pobierają dwutlenek węgla, który w cytoplazmie komórek miękiszu asymilacyjnego jest przyłączany do związku trójwęglowego – fosfoenolopirogronianu (PEP). W konsekwencji powstaje związek czterowęglowy – szczawiooctan (OAA). Następnie szczawiooctan ulega redukcji do jabłczanu, który gromadzony jest w wakuoli. Zdolność do gromadzenia jabłczanu (kwasu jabłkowego) dało podstawy do sformułowania nazwy tej grupy roślin – CAM (z ang. crassulacean acid metabolism). W dzień aparaty szparkowe roślin typu CAM są zamknięte. Następuje wówczas transport jabłczanu z wakuoli do cytoplazmy. W cytoplazmie komórek miękiszu asymilacyjnego następuje odłączenie dwutlenku węgla od jabłczanu, co skutkuje powstaniem związku trójwęglowego – pirogronianu, z którego zostanie odtworzony fosfoenolopirogronian (PEP). Uwolniony dwutlenek węgla zostaje przyłączony do rybulozo‑1,5‑bisfosforanu (RuBP) i podlega przemianom cyklu Calvina, tak samo jak w fotosyntezie typu CIndeks dolny 33.
Podobieństwa i różnice między fotosyntezą typu CIndeks dolny 44 i CAM
W ogólnym ujęciu przebieg fotosyntezy CIndeks dolny 44 i CAM jest podobny. W obu szlakach fotosyntetycznych ma miejsce mechanizm dwuetapowej asymilacji dwutlenku węgla. Różnice między fotosyntezą typu CIndeks dolny 44 i CAM dotyczą czasu oraz miejsca asymilacji dwutlenku węgla i dekarboksylacjidekarboksylacji jabłczanu.
Rośliny typu CIndeks dolny 44:
wiążą COIndeks dolny 22 do fosfoenolopirogronianu w dzień, w komórkach miękiszu asymilacyjnegomiękiszu asymilacyjnego (mezofilu);
powstały związek czterowęglowy ulega dekarboksylacji w komórkach pochwy okołowiązkowej.
Zatem: procesy przyłączania COIndeks dolny 22 do związku trójwęglowego i odłączania COIndeks dolny 22 od związku czterowęglowego są rozdzielone przestrzennie, gdyż zachodzą w różnych komórkach.
Rośliny typu CAM:
przyłączają COIndeks dolny 22 do fosfoenolopirogronianu w nocy, w komórkach miękiszu asymilacyjnego;
powstały związek czterowęglowy ulega dekarboksylacji w dzień w tych samych komórkach co uprzednio, tzn. w komórkach miękiszu asymilacyjnego.
Zatem: procesy przyłączania COIndeks dolny 22 do związku trójwęglowego i odłączania COIndeks dolny 22 od związku czterowęglowego są rozdzielone czasowo, gdyż zachodzą w różnych porach w obrębie tej samej komórki.
Porównanie fotosyntezy typu CIndeks dolny 33, CIndeks dolny 44 i CAM
Cechy porównywane | Rodzaj fotosyntezy | ||
|---|---|---|---|
CIndeks dolny 33 | CIndeks dolny 44 | CAM | |
Etapy asymilacji | jeden etap | dwa etapy – rozdzielone przestrzennie | dwa etapy – rozdzielone czasowo |
Miejsce asymilacji | miękisz asymilacyjny | etap – miękisz asymilacyjny | miękisz asymilacyjny |
etap 2 – komórki pochwy okołowiązkowej | |||
Pora asymilacji | dzień | dzień | etap 1 – noc |
etap 2 – dzień | |||
Enzym katalizujący reakcję asymilacji | karboksylaza rybulozo‑1,5‑bisfosforanu (RuBisCO) | etap 1 – karboksylaza fosfoenolopirogronianowa (PEPC) | etap 1 – karboksylaza fosfoenolopirogronianowa (PEPC) |
etap 2 – karboksylaza rybulozo‑1,5‑bisfosforanu (RuBisCO) | etap 2 – karboksylaza rybulozo‑1,5‑bisfosforanu (RuBisCO) | ||
Pierwszy akceptor COIndeks dolny 22 | rybulozo‑1,5‑bisfosforan (RuBP) | fosfoenolopirogronian (PEP) | fosfoenolopirogronian (PEP) |
Pierwszy produkt karboksylacji | kwas 3‑fosfoglicerynowy (PGA) | szczawiooctan (OAA) | szczawiooctan (OAA) |
Słownik
inaczej przyswajanie; proces przekształcania substancji pobranych z otoczenia dla potrzeb metabolicznych komórki
rodzaj reakcji chemicznej polegającej na usunięciu grupy karboksylowej (COOH), np. z cząsteczki kwasu karboksylowego, w wyniku czego dochodzi do wydzielenia dwutlenku węgla
inaczej cykl Calvina lub cykl Calvina‑Bensona; cykl biochemiczny zachodzący w stromie chloroplastów w komórkach roślin; faza ciemna fotosyntezy – niezależna od światła, ale wykorzystująca produkty fazy jasnej fotosyntezy: ATP i NADPH; dzieli się na trzy etapy: karboksylacji, redukcji i regeneracji; polega na przekształceniu dwutlenku węgla w glukozę
jedna z dwóch faz fotosyntezy; faza zależna od światła zachodząca w błonach tylakoidów gran i polegająca na przekształceniu energii świetlnej w energię wiązań chemicznych; pozwala na wytworzenie związków wykorzystywanych w fazie ciemnej fotosyntezy: ATP i NADPH
rodzaj reakcji chemicznej polegającej na przyłączeniu dwutlenku węgla do innego związku chemicznego, w wyniku czego powstaje kwas karboksylowy
rodzaj tkanki miękiszowej, której komórki zawierają chloroplasty; tkanka odpowiedzialna za przeprowadzanie procesu fotosyntezy; u większości roślin obecna w liściach, rzadziej występuje w łodydze lub korzeniach; zazwyczaj tkanka zróżnicowana na miękisz palisadowy i gąbczasty
rodzaj połączenia pomiędzy komórkami roślinnymi; tworzone przez cienkie pasma cytoplazmy przenikające przez jamki w ścianach komórkowych sąsiadujących ze sobą komórek; umożliwiają wymianę substancji między komórkami
fizjologiczny proces parowania wody z powierzchni roślin; zachodzi głównie przez aparaty szparkowe liści, a w niewielkim stopniu także przez kutykulę; powoduje powstawanie siły ssącej liści umożliwiającej podciąganie wody z korzenia w górę rośliny