Przeczytaj
Do lat 40. XX wieku zakładano, że materiał genetyczny ma skomplikowaną budowę. W końcu – myśleli uwcześni naukowcy – warunkuje on funkcjonowanie niezwykle złożonych organizmów i zapewnia dziedziczenie ogromnej liczby cech. Przypuszczano więc, że rolę materiału genetycznego pełnią białka – wielkocząsteczkowe związki organiczne o niezwykle skomplikowanej budowie. Białka zbudowane są z dwudziestu różnych aminokwasów, które łączą się ze sobą, tworząc złożone struktury przestrzenne. Dla porównania kwasy nukleinowe – inne znane już wówczas związki wielkocząsteczkowe – zbudowane są z czterech rodzajów nukleotydów. Nukleotydy łączą się ze sobą w regularny sposób i tworzą stosunkowo prostą strukturę przestrzenną. Tak wyraźny kontrast pomiędzy strukturą białek i kwasów nukleinowych skłaniał ówczesnych badaczy do błędnego przekonania, że materiałem genetycznym są białka. Dopiero dalsze badania nad mikroorganizmami dostarczyły pierwszych dowodów, że nośnikiem informacji genetycznej jest DNA.
Doświadczenie Griffitha
W 1928 roku Frederick Griffith przeprowadził doświadczenie, w którym wykorzystał dwa szczepyszczepy dwoinki zapalenia płuc Streptococcus pneumoniae:
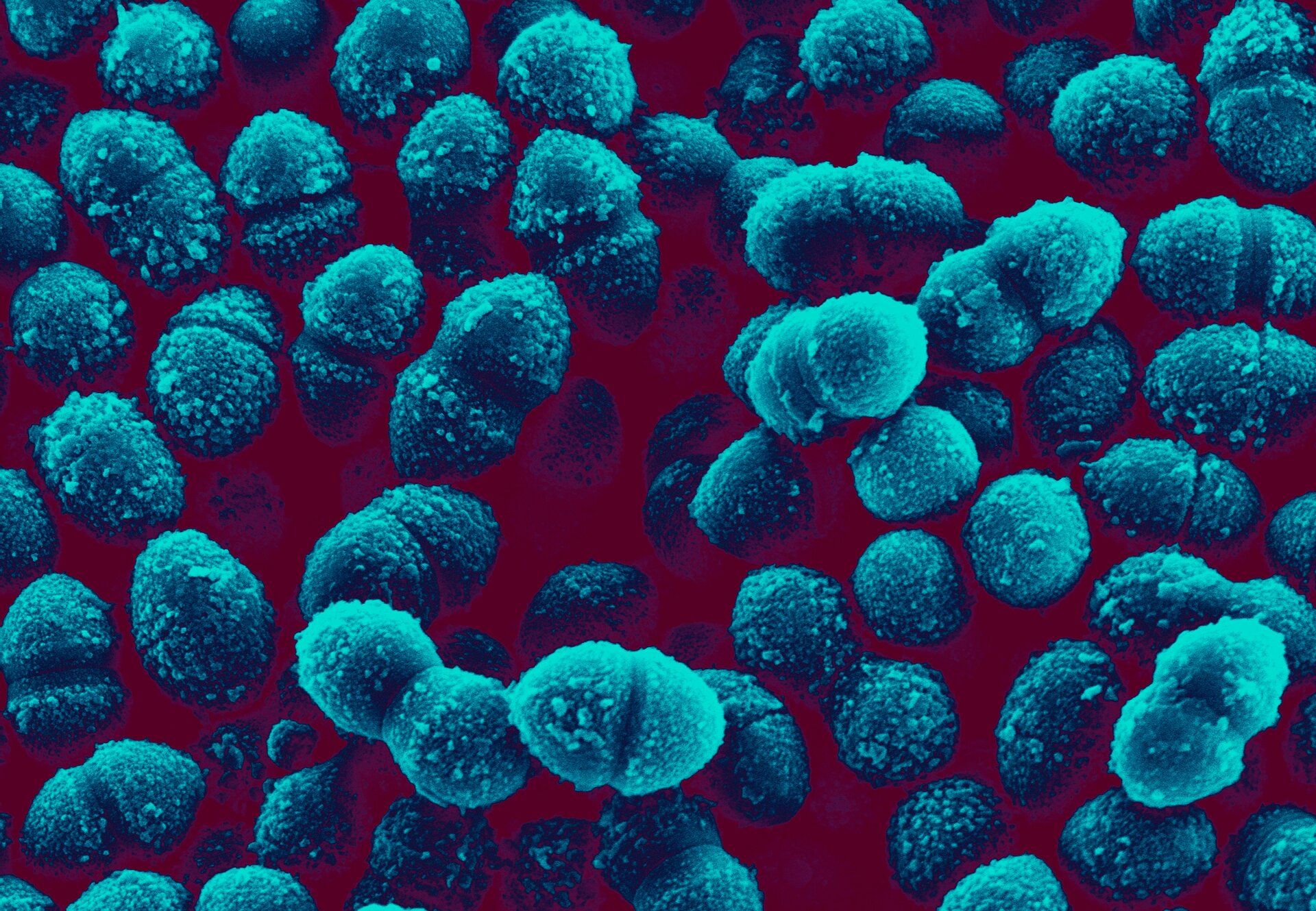
Na zewnątrz ściany komórkowej bakterii chorobotwórczego szczepu S znajduje się polisacharydowa otoczka, która skutecznie chroni bakterie przed układem odpornościowym zainfekowanego organizmu. Bakterie szczepu R nie mają tej otoczki, przez co nie są chorobotwórcze. Zdjęcie spod skaningowego mikroskopu elektronowego, koloryzowane, powiększenie 100×.
W czasie eksperymentu zwierzętom laboratoryjnym wstrzykiwano różne roztwory komórek bakterii Streptococcus pneumoniae. Podanie bakterii szczepu S wywoływało zapalenie płuc. Natomiast podanie bakterii szczepu R nie prowadziło do rozwoju choroby. Następnie roztwór ze zjadliwym szczepem S podgrzano w wysokiej temperaturze, co unieszkodliwiło komórki bakterii. Podanie martwych bakterii szczepu S nie skutkowało wystąpieniem objawów choroby.
Podanie mieszaniny martwych bakterii szczepu S i żywych bakterii szczepu R prowadziło do wystąpienia zapalenia płuc i w konsekwencji śmierci zwierząt laboratoryjnych. W próbkach krwi pobranych od martwych myszy wykryto żywe bakterie należące do szczepu S. Żywe i niechorobotwórcze komórki bakterii szczepu R pobrały pewien czynnik z martwych komórek szczepu S, w wyniku czego przekształciły się w formy chorobotwórcze. Jednak czynnik, który spowodował transformacjętransformację komórek bakterii, pozostawał nieznany.

Wyniki uzyskane przez Griffitha były przesłanką do odrzucenia dotychczasowego przekonania, że to białka stanowią nośnik informacji genetycznej. Wciąż jednak brakowało ostatecznego wyjaśnienia, jakim związkiem chemicznym był czynnik transformujący bakterie.
Doświadczenie Avery’ego
Dopiero kilkanaście lat po doświadczeniu Griffitha udało się określić chemiczne właściwości czynnika transformującego bakterie Streptococcus pneumoniae. W 1944 roku Oswald Avery, Colin MacLeod oraz Maclyn McCarty przeprowadzili doświadczenie, w którym wykorzystali unieszkodliwione pod wpływem wysokiej temperatury komórki bakterii szczepu S.
Martwe bakterie potraktowano detergentem, który doprowadził do lizylizy komórek. Uzyskany materiał odwirowano, aby oddzielić zawartość komórki od pozostałości ściany komórkowej i polisacharydowej otoczki. Otrzymano ekstrakt komórkowy zdolny do transformacji żywych komórek bakterii szczepu R.
Aby sprawdzić, jaka substancja chemiczna jest „czynnikiem transformującym” odkrytym przez Griffitha, do ekstraktu komórkowego kolejno dodawano proteazę, RNAzę i DNAzę, czyli enzymy degradujące odpowiednio białka, RNA oraz DNA. Tylko w próbie, w której DNA został rozłożony przez DNAzę, ekstrakt komórkowy stracił zdolność transformacji bakterii szczepu R.
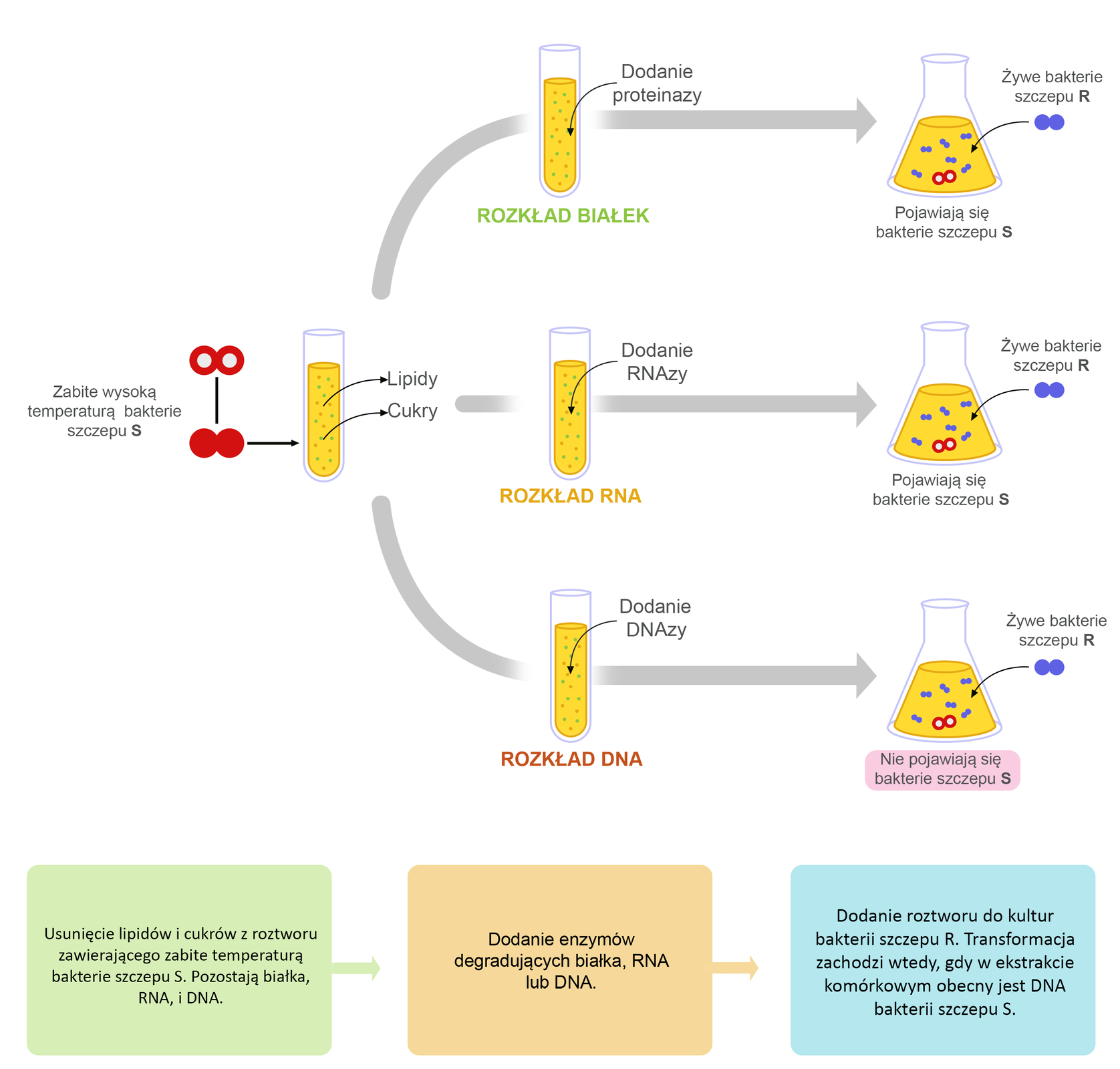
Wyniki uzyskane przez Avery’ego i współpracowników dowiodły, że czynnikiem modyfikującym informację genetyczną badanych bakterii jest DNA.
Doświadczenie Hersheya i Chase

Późniejsze doświadczenie opisujące cykl namnażania się bakteriofagówbakteriofagów pozwoliło jednoznacznie określić rolę DNA jako nośnika informacji genetycznej. W 1952 roku Alfred Hershey i Martha Chase przeprowadzili doświadczenie, w którym wykorzystali bakteriofaga T2 infekującego komórki bakterii – pałeczki okrężnicy (Escherichia coli). Celem ich pracy było określenie, który z dwóch komponentów tworzących bakteriofagi wnika do komórek gospodarza i umożliwia wytworzenie nowych wirionówwirionów.
Białkowy kapsyd bakteriofagów zawiera atomy siarki w postaci grup tiolowychgrup tiolowych (grupy te wchodzą w skład niektórych aminokwasów), jednak nie ma w nim atomów fosforu. Z kolei DNA zawiera atomy fosforu (w postaci reszt fosforanowych budujących nukleotydy), lecz jest pozbawiony atomów siarki.
W celu rozróżnienia białkowego kapsydu i DNA wirusa wykorzystano radioaktywne izotopy siarki i fosforu. W pierwszej fazie doświadczenia część bakterii hodowano na pożywce zawierającej radioaktywną siarkę (Indeks górny 3535S), a część na pożywce zawierającej radioaktywny fosfor (Indeks górny 3232P). Obie grupy bakterii wykorzystały izotopy siarki i fosforu do budowy własnych białek i DNA.
Następnie bakterie infekowano bakteriofagami T2. Wirusy namnażały się, korzystając ze zdolności metabolicznych komórek bakterii. W efekcie uzyskano dwie grupy bakteriofagów: pierwsza zawierała kapsyd z wbudowanym izotopem Indeks górny 3535S, a druga DNA z wbudowanym izotopem Indeks górny 3232P.
W drugiej fazie doświadczenia bakterie hodowane na zwykłej pożywce, niezawierającej promieniotwórczych izotopów siarki ani fosforu, infekowano wyznakowanymi bakteriofagami. W pierwszej próbie wykorzystano bakteriofagi, których białkowy kapsyd zawierał izotop Indeks górny 3535S. W drugiej próbie znajdowały się bakteriofagi zawierające izotop Indeks górny 3232P w wirusowym DNA.
Po pewnym czasie każdą z prób zmiksowano, aby odłączyć białkowe kapsydy bakteriofagów od bakterii. Następnie uzyskane mieszaniny poddano wirowaniu w celu rozdzielenia roztworu na dwie frakcje. W ten sposób otrzymano osad zawierający komórki bakterii i supernatantsupernatant (nadsącz), w którym znajdowały się białkowe kapsydy bakteriofagów.
W pierwszej próbie, znakowanej izotopem siarki Indeks górny 3535S, radioaktywny znacznik znajdował się w supernatancie. Pozwoliło to stwierdzić, że białkowy kapsyd nie wnika do wnętrza bakterii w czasie infekcji.
W drugiej próbie radioaktywny izotop Indeks górny 3232P znajdował się w osadzie. W ten sposób dowiedziono, że w czasie infekcji bakteriofagii wstrzykują do wnętrza komórek bakterii wirusowy DNA.
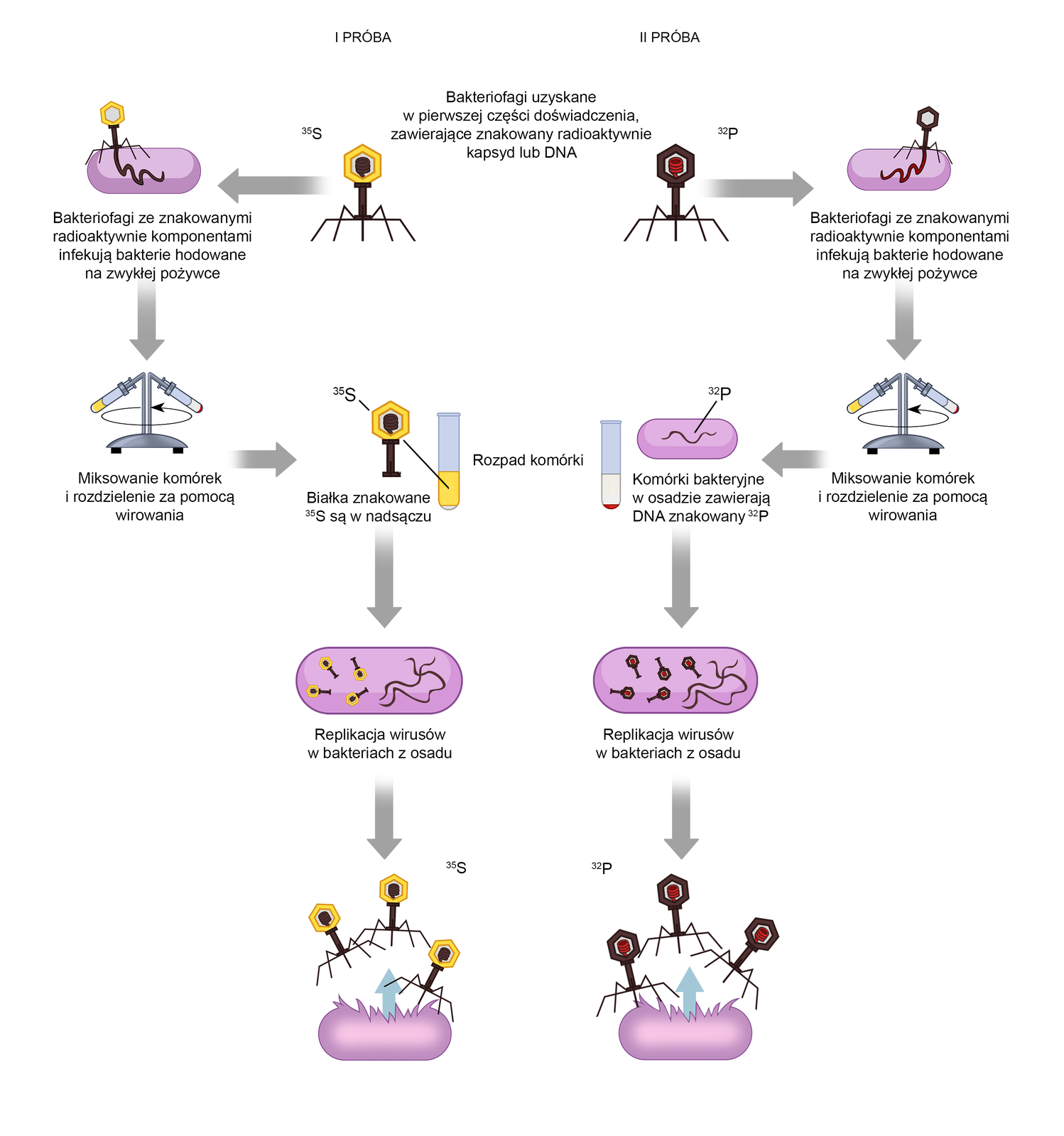
Wyniki uzyskane przez Hersheya i Chase dowiodły, że nośnikiem informacji genetycznej bakteriofagów jest DNA, a nie białko.
Słownik
(gr. bakteria – bakteria, phage – jeść) wirusy atakujące wyłącznie bakterie; zbudowane z białkowego kapsydu składającego się z główki, ogonka i włókienek, wewnątrz którego obecny jest materiał genetyczny w postaci DNA lub rzadziej RNA; wirusy te są swoiste względem gatunku lub szczepu bakterii
(−SH); grupa sulfohydrylowa, analogiczna do grupy wodorotlenowej (-OH); występuje w tiolach oraz cysteinie (jeden z aminokwasów)
(gr. lysis – rozpad) rozpad komórki na skutek przerwania ciągłości jej błony komórkowej
roztwór pozostały po opadnięciu osadu z mieszaniny
czysta hodowla bakterii danego gatunku, pochodząca od pojedynczej komórki, mająca unikalny genotyp i fenotyp
(łac. transformatio) zjawisko pobierania przez bakterie materiału genetycznego z podłoża lub jego wymiana między komórkami bakteryjnymi
pojedyncza, kompletna cząstka wirusa zdolna do infekowania komórek bakteryjnych, roślinnych i zwierzęcych
inaczej wirulencja; zdolność patogenów do wniknięcia, namnażania się oraz wywołania choroby w zainfekowanym organizmie