Ekologia populacji i ekosystemów
Zależności antagonistyczne
Scharakteryzujesz poszczególne rodzaje oddziaływań antagonistycznych.
Wskażesz przykłady gatunków, między którymi występują oddziaływania antagonistyczne.
Przeprowadzisz doświadczenie wykazujące interakcję antagonistyczną między organizmami należącymi do dwóch różnych gatunków pantofelków (Paramecium).
Odziaływania antagonistyczneantagonistyczne między organizmami przynoszą straty co najmniej jednej ze stron uczestniczących w interakcji. Niekorzystny wpływ jednego organizmu na drugi najczęściej dotyczy ograniczenia: liczebności, przeżywalności i rozrodczości.
Oddziaływania antagonistyczne to zależności między dwoma organizmami, najczęściej należącymi do różnych gatunków, z których co najmniej jeden ponosi straty. Do takich interakcji zalicza się: drapieżnictwo, roślinożerność, pasożytnictwo, konkurencję, allelopatię i amensalizm.
Drapieżnictwo
Drapieżnictwo to rodzaj oddziaływania antagonistycznego, w którym drapieżnik aktywnie poluje na ofiarę, zabija ją i zjada. Organizmem odnoszącym korzyści jest drapieżnik, natomiast ofiara to organizm ponoszący straty. Poprzez swoje działanie drapieżnik dokonuje selekcji – wśród potencjalnych ofiar dochodzi do wyeliminowania osobników najsłabszych, np. młodych, starych, chorych lub okaleczonych. W środowisku pozostają osobniki najlepiej przystosowane do życia, a ich przestrzeń życiowa i dostęp do zasobów środowiska się zwiększają. Ponadto mniejsza liczebność populacji ofiary jednego gatunku sprawia, że w środowisku dostępna jest większa ilość pokarmu dla populacji ofiary drugiego gatunku. Zatem drapieżniki pełnią funkcję regulacyjną, zmniejszając napięcia powstające na skutek konkurencji wewnątrzgatunkowej i międzygatunkowej.

Przystosowania drapieżników do polowania
Przystosowania drapieżników mają na celu zwiększenie prawdopodobieństwa znalezienia, skutecznego zaatakowania i uśmiercenia potencjalnej ofiary. Drapieżnictwo jest możliwe dzięki przystosowaniom: morfologicznymmorfologicznym, anatomicznoanatomiczno-fizjologicznymfizjologicznym, chemicznym i behawioralnym.
Drapieżniki mogą mieć ubarwienie maskujące, które umożliwia skradanie się i prawie niezauważalne podejście do potencjalnej ofiary.

W budowie morfologicznej drapieżników zaznacza się obecność różnego rodzaju struktur umożliwiających schwytanie ofiary i jednocześnie zmniejszających szansę upolowanego zwierzęcia na uwolnienie się. Należą do nich: odnóża chwytne, szpony i pazury.


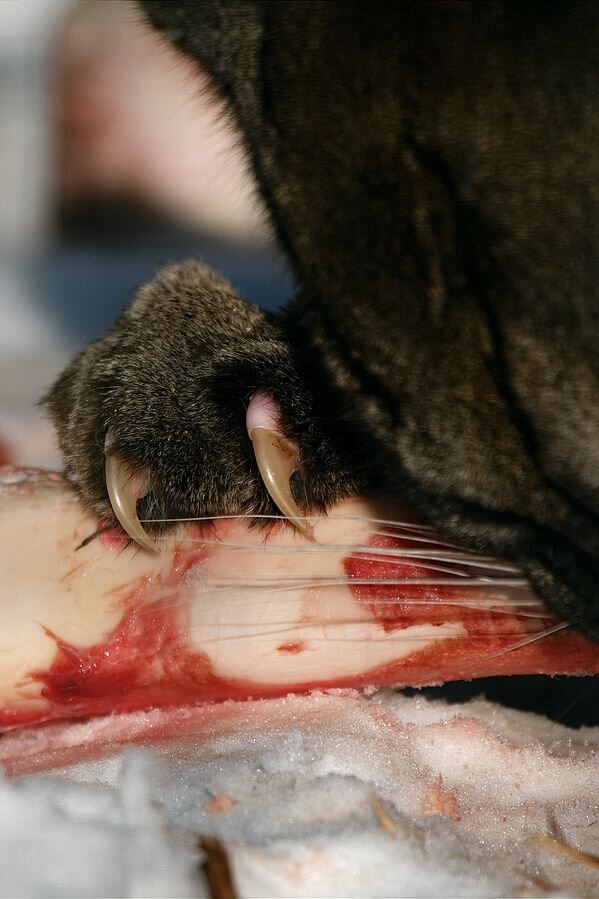
Budowa anatomiczna i fizjologia drapieżników jest wyrazem ich przystosowania do polowań. Szereg adaptacji dotyczy budowy i funkcjonowania układu pokarmowego oraz układu mięśniowego.
Układ pokarmowy
U drapieżników początkowe odcinki układu pokarmowego zaopatrzone są w struktury umożliwiające schwytanie ofiary oraz jej zabicie. Takimi strukturami są m.in.: aparat gębowy typu gryzącego, szczękoczułki i nogogłaszczki, zęby, długi i lepki język
Układ mięśniowy
Mięśnie drapieżników są silne i dobrze rozwinięte, co umożliwia aktywny atak, pogoń za uciekającą ofiarą i jej zabicie.

Niektóre drapieżniki budują pułapki, w które wpada potencjalna ofiara.


Drapieżniki są zdolne do produkcji i wydzielania substancji chemicznych o różnorodnym działaniu. Najczęściej substancje te są silnie toksyczne, a po wprowadzeniu do ciała ofiary uszkadzają jej układ nerwowy, zazwyczaj prowadząc do paraliżu.


Przystosowania behawioralne drapieżników obejmują różnego rodzaju techniki łowieckie, wykorzystywane w czasie polowania. Techniki polowania obejmują: tropienie, skradanie się i pościg, atak z ukrycia lub atak z zaskoczenia. Szanse na skuteczne upolowanie potencjalnej ofiary rosną, jeśli osobniki tworzące stado współpracują ze sobą.


Przystosowania ofiar do obrony przed drapieżnikami
Przystosowania ofiar do obrony mają na celu zmniejszenie prawdopodobieństwa skutecznego ataku, odstraszenie lub zniechęcenie drapieżnika. Mechanizmy obronne są możliwe dzięki przystosowaniom: morfologicznym, anatomiczno‑fizjologicznym, chemicznym i behawioralnym.
W budowie morfologicznej potencjalne ofiary wykształcają różnego rodzaju struktury ochronne, które stanowią element obrony mechanicznej przed atakiem ze strony drapieżnika. Strukturami ochronnymi są: domki, muszle, kolce, poroże i rogi.
Elementem budowy morfologicznej zmniejszającym możliwość dostrzeżenia ofiary i tym samym ograniczającym ryzyko ataku ze strony drapieżnika jest ubarwienie ochronne. Odpowiednie ubarwienie może być wynikiem mimetyzmumimetyzmu lub mimikrymimikry.


Potencjalne ofiary mogą mieć bardzo dobrze rozwinięte zmysły, które wykorzystują w celu zapewnienia sobie bezpieczeństwa. Bardzo dobrze rozwinięty słuch, węch i wzrok umożliwiają stałą kontrolę otoczenia i pozwalają wyczuć zagrożenie w odpowiednim momencie, gdy jest jeszcze czas na ucieczkę.

Rozpoznanie zagrożenia ze strony drapieżnika sprawia, że potencjalne ofiary podejmują obronę w formie ucieczki. Skuteczność tej metody zależy od uwarunkowań morfologicznych i fizjologicznych.

Potencjalne ofiary są zdolne do produkcji i wydzielania substancji o działaniu toksycznym lub drażniącym. Niektóre zwierzęta mają w skórze gruczoły jadowe produkujące jad; wówczas atak drapieżnika zazwyczaj kończy się wypluciem ofiary. Inne zwierzęta w sytuacji zagrożenia wydzielają do otoczenia substancje mające na celu odstraszenie i zniechęcenie drapieżnika do kontynuowania ataku.


Obronnymi przygotowaniami behawioralnymi są różnego rodzaju zachowania potencjalnych ofiar. Należą się do nich: autotomiaautotomia, życie w stadzie, wydzielanie substancji chemicznych.




Aby rozwiązać polecenia przeanalizuj poniższe grafiki o rodzajach mimikry.
Mimikra polega na upodobnieniu się do innych zwierząt, które ze względu na swą jadowitość, trujący lub odrażający smak nie są atakowane przez drapieżniki.
Mimikra batesowska (Batesa, batezjańska) dotyczy sytuacji, w której bezbronny gatunek upodabnia się do gatunku mogącego się bronić dzięki obecności struktur obronnych: kolców, żądła, gruczołów jadowych.
Mimikra müllerowska dotyczy sytuacji, w której dwa gatunki mogące się bronić upodabniają się do siebie.
Układ drapieżca–ofiara
Duża liczba potencjalnych ofiar sprzyja zwiększeniu się populacji drapieżnika – w ten sposób regulowana jest liczebność obu gatunków na zamieszkiwanym przez nie obszarze. Jeśli w wyniku intensywnych polowań populacja ofiar ulegnie zmniejszeniu, w konsekwencji, po pewnym czasie, zmniejszy się też liczba drapieżców – w środowisku będzie dla nich za mało pożywienia. I odwrotnie: zmniejszenie liczebności drapieżników wpłynie na odrodzenie się populacji ofiar. To przykład na cykliczność zmian liczebności w obu populacjach – taka regulacja ma charakter ujemnego sprzężenia zwrotnego: mniej pokarmu – mniej drapieżników, mniej drapieżników – więcej ich ofiar, czyli pokarmu.
W zróżnicowanym gatunkowo środowisku naturalnym drapieżnik zaprzestaje polowań na osobniki zmniejszonej populacji, bo w zasięgu jego węchu i wzroku pojawiają się inne, łatwo dostępne potencjalne ofiary.
Szczyt liczebności drapieżników występuje nieco później niż ich ofiar. Obfitość dostępnego pokarmu sprzyja rozrodowi i przeżywaniu potomstwa drapieżców, ale wzrost populacji zaczyna się wtedy, gdy młode drapieżniki same osiągną wiek rozrodu. Populacja ofiar jest już w tym momencie zmniejszona, co jest wynikiem wcześniejszego zwiększenia intensywności polowań przez drapieżniki w związku z koniecznością wykarmienia większej liczby potomstwa.

Przykładem zależności między liczebnością populacji drapieżników i ofiar są zbadane cykliczne wahania liczebności populacji zająca bielaka i rysia kanadyjskiego na terenie Kanady. Wpływa na nie np. dostępność pokarmu dla roślinożerców. Stwierdzono, że intensywne podgryzanie roślin przez liczną populację zajęcy powodowało odkładanie się w młodych gałązkach trujących związków fenolowych. Następstwem tego zjawiska była rosnąca śmiertelność zajęcy. Rośliny, które nie były już podgryzane, po pewnym czasie wzrostu bez przeszkód nie odkładały już toksycznych związków. Po kilku latach liczba zajęcy mogła ponownie szybko wzrosnąć – wraz z nią wzrastała liczba polujących na nie rysi. Cykl ten powtarzał się regularnie przez wiele lat.


Rysie, największe drapieżne koty w Polsce, występują na obszarze około 12% polskich lasów – żyje w nich zaledwie ok. 200 osobników. To zdecydowanie za mało, aby gatunek mógł przetrwać bez pomocy człowieka, dlatego podlega ścisłej ochronie.
Rysie nie lubią otwartych przestrzeni. Bytują głównie we wschodnich i południowych częściach kraju, gdzie lasy są najbardziej rozległe. Odżywiają się ptakami, drobnymi gryzoniami, zającami i królikami – gdy ich brakuje, szczególnie w zimie, polują na sarny i jelenie.
Dużym zagrożeniem dla rysi jest nieprawidłowo prowadzona gospodarka leśna, np. nadmierny odstrzał zajęcy i saren.

Przeprowadź symulację zmian liczebności populacji drapieżnika i ofiary. Następnie odpowiedz na polecenia.
Kliknij na poszczególne elementy symulacji i obserwuj, jak zmienia się liczebność zależnych od siebie populacji – zająca bielaka (Lepus timidus) i rysia kanadyjskiego (Lynx canadensis) na przestrzeni lat.
Interaktywny wykres liniowy przedstawia zmiany liczebności dwóch populacji w latach 1845–1935: zająca bielaka oraz rysia kanadyjskiego. Oś X to lata (co 5 lat), oś Y to liczebność populacji (wartości od kilku tysięcy do 160 tysięcy). Na wykresie są dwie linie – osobno dla zająca i rysia – co umożliwia porównanie przebiegu zmian w czasie.
Populacja zająca zmienia się bardzo silnie: startuje od 20 000 (1845), rośnie do około 60 000 (1850–1855), spada do 22 000 (1860), po czym osiąga najwyższą wartość 160 000 (1865). Następnie gwałtownie spada do 10 000 (1870) i 8 000 (1880). Kolejny duży wzrost to 100 000 (1875) oraz 140 000 (1885). W późniejszym okresie zając osiąga m.in. 80 000 (1895), spada do 6 000 (1900), rośnie do 75 000 (1905), spada do 30 000 (1910), a minimum wynosi 5 000 (1920). Na końcu serii wartości wynoszą 70 000 (1925), 20 000 (1930) i 76 000 (1935).
Populacja rysia również faluje, ale w mniejszym zakresie: w 1845 wynosi 60 000, spada do 20 000 (1850), rośnie do 30 000 (1855), spada do 10 000 (1860) i rośnie do 70 000 (1865). Później pojawiają się niższe wartości 10 000 (1870 i 1880), wzrost do 40 000 (1875) oraz najwyższa wartość 80 000 (1885). Kolejne zmiany to 19 000 (1890), 50 000 (1895), minimum 4 000 (1900), wysoka wartość 70 000 (1905), spadek do 10 000 (1910), wzrost do 50 000 (1915), 5 000 (1920), 50 000 (1925), 20 000 (1930) i 38 000 (1935).
Wykres pozwala obserwować naprzemienne wzrosty i spadki obu populacji oraz porównać ich wartości w tych samych latach.
Roślinożerność
Roślinożerność to rodzaj oddziaływania antagonistycznego, w którym roślinożerca zjada zazwyczaj tylko część rośliny. Organizmem odnoszącym korzyści jest roślinożerca, natomiast roślina ponosi straty. Uszkodzone rośliny najczęściej mają możliwość dalszego wzrostu, jednak osłabione mogą przegrać konkurencję z innymi roślinami lub stać się celem ataku ze strony pasożytów. Bardzo rzadko, w czasie masowego pojawienia się roślinożerców, może dojść do całkowitego wyniszczenia populacji roślin.
Pokarm roślinny jest bardzo zróżnicowany. Zalicza się do niego liście, owoce, trawę, korę drzew, nasiona, nektar, pyłek i glony. Ze względu na spożywane części roślin wyróżnia się różne rodzaje roślinożerców - fitofagów.
Adaptacje roślinożerców do zdobywania pokarmu
Roślinożercy przystosowali się do zdobywania pokarmu dzięki adaptacjom anatomicznych i behawioralnym.
Adaptacje anatomiczne roślinożerców do zjadania roślin dotyczą przede wszystkim:
budowy zębów,
budowy przewodu pokarmowego.



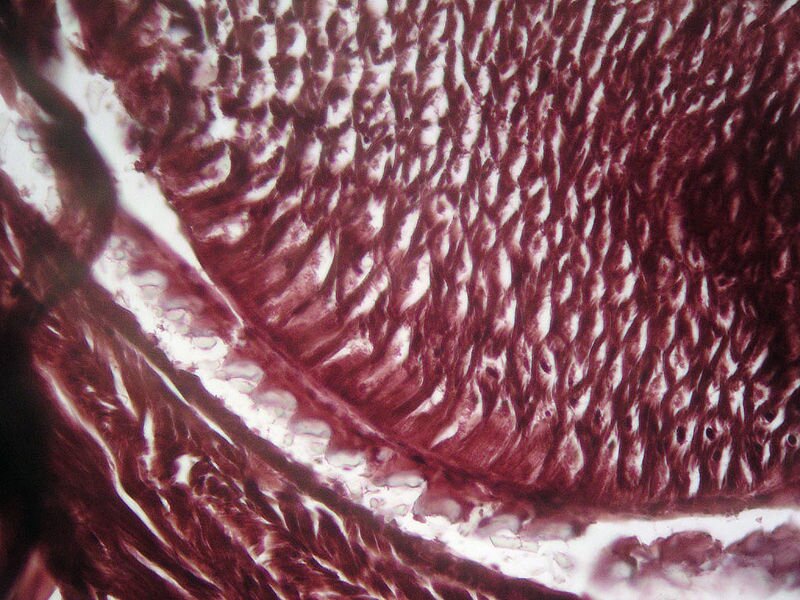





Adaptacje behawioralne roślinożerców do zjadania roślin obejmują przede wszystkim:
umiejętność odróżniania roślin jadalnych od roślin trujących;
umiejętność odróżniania jadalnych części roślin od trujących części roślin;
dopasowanie swojego cyklu życiowego do czasu występowania trujących substancji w roślinie.
Przeanalizuj poniższe grafiki, a później odpowiedz na polecenia.
Najbardziej zaawansowane przystosowania do trawienia pokarmu roślinnego wykształciły przeżuwacze, do których zalicza się bydło, owce, jelenie, żyrafy, łosie i żubry.
Mechanizmy obronne przed roślinożercami
Wiele roślin wykształciło mechanizmy obronne chroniące je przed roślinożercami. Wyróżniamy trzy rodzaje tych mechanizmów:
Wiele gatunków roślin produkuje różne związki chemiczne o działaniu trującym lub odstraszającym zwierzęta. Mogą się one znajdować w owocach, liściach, pędach lub nasionach. Inna strategia obronna polega na wytwarzaniu substancji, które nadają roślinie nieprzyjemny smak lub zapach. Jest on warunkowany obecnością tanintanin.







Obrona mechaniczna polega na wytwarzaniu grubej warstwy tkanki sklerenchymatycznej, która utrudnia zgryzanie liści i łodyg lub na wykształceniu ciernicierni, kolcówkolców lub włoskówwłosków parzących.




U roślin obrona bierna polega na upodabnianiu się całej lub części rośliny do otoczenia lub gatunków unikanych przez roślinożerców. Takie zjawisko to mimetyzm.



Inna roślina posiadająca włoski parzące to barszcz Sosnowskiego (Heracleum sosnovskyi). Jego pęd pokryty jest włoskami uwalniającymi furanokumarynę, która powoduje oparzenia chemiczne tkanek miękkich zwierząt. Włoski z tą substancją unoszą się również w powietrzu, w bezpośrednim sąsiedztwie rośliny.
Gatunek jest niebezpieczny dla zdrowia i życia ludzi oraz zwierząt hodowlanych.

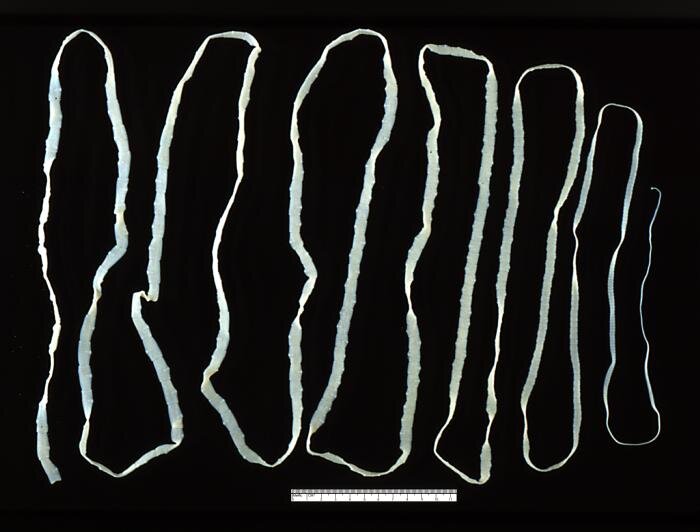
Pasożytnictwo
Pasożytnictwo to rodzaj oddziaływania antagonistycznego, w którym pasożyt pobiera substancje odżywcze od żywiciela i często wykorzystuje go również jako środowisko życia. Organizmem odnoszącym korzyści jest pasożyt, natomiast żywiciel ponosi straty. Zazwyczaj pasożyt nie doprowadza do śmierci żywiciela. Jednak jego obecność osłabia gospodarza, który staje się łatwiejszym celem dla drapieżnika lub przegrywa konkurencję z innymi organizmami.

Przystosowania pasożytów do prowadzenia pasożytniczego trybu życia
Przystosowania do pasożytnictwa mają na celu znalezienie żywiciela i jego skuteczne zakażenie oraz rozmnożenie pasożyta i wydanie na świat licznego potomstwa, Pozwoli to rozprzestrzenić gatunek w środowisku i jednocześnie zwiększy prawdopodobieństwo znalezienia odpowiedniego organizmu żywicielskiego. Prowadzenie pasożytniczego trybu życia jest możliwe dzięki przystosowaniom: morfologicznym, anatomiczno‑fizjologicznym, chemicznym i behawioralnym.
Budowa morfologiczna pasożytów jest wyrazem przystosowania do prowadzenia pasożytniczego trybu życia. Kształt ich ciała wiąże się ze środowiskiem życia i sposobem pobierania pokarmu. Zazwyczaj jest ono spłaszczone grzbietobrzusznie, rzadziej bocznie, może być też obłe.


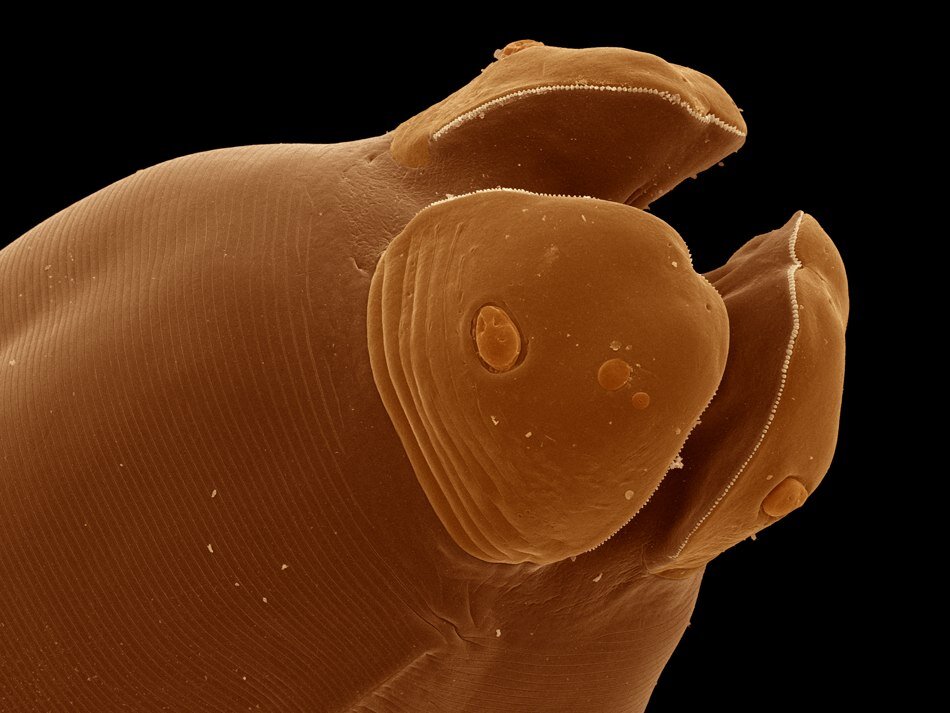
W budowie morfologicznej pasożytów obecne są różnego rodzaju struktury umożliwiające przyczepienie się do ciała żywiciela. Występowanie pazurków, haczyków, przyssawek, bruzd, ząbków i sztylecików zapewnia umocowanie pasożyta i jednocześnie chroni go przed usunięciem z powierzchni ciała żywiciela lub z jego wnętrza.

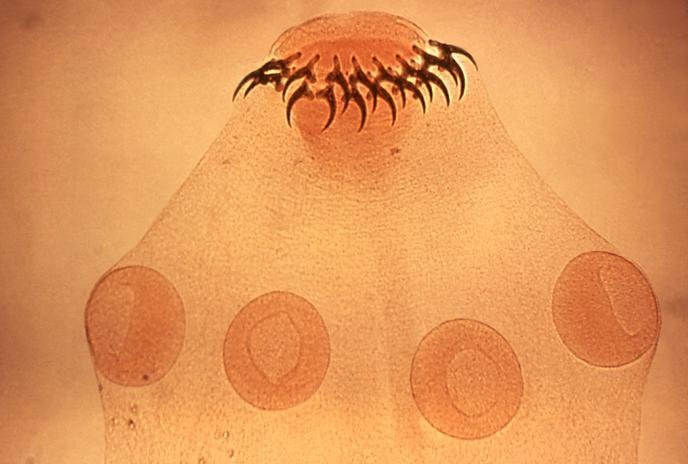
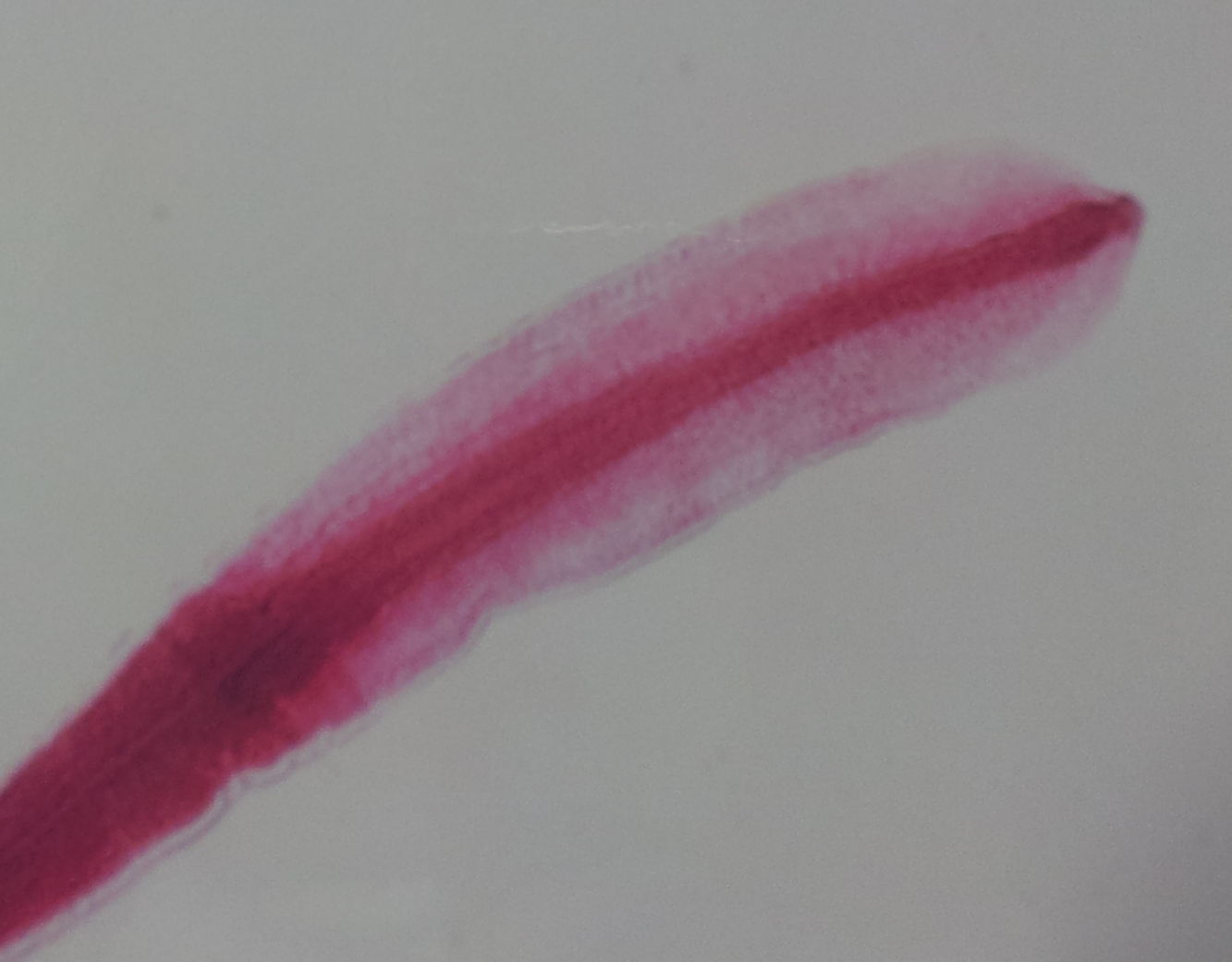
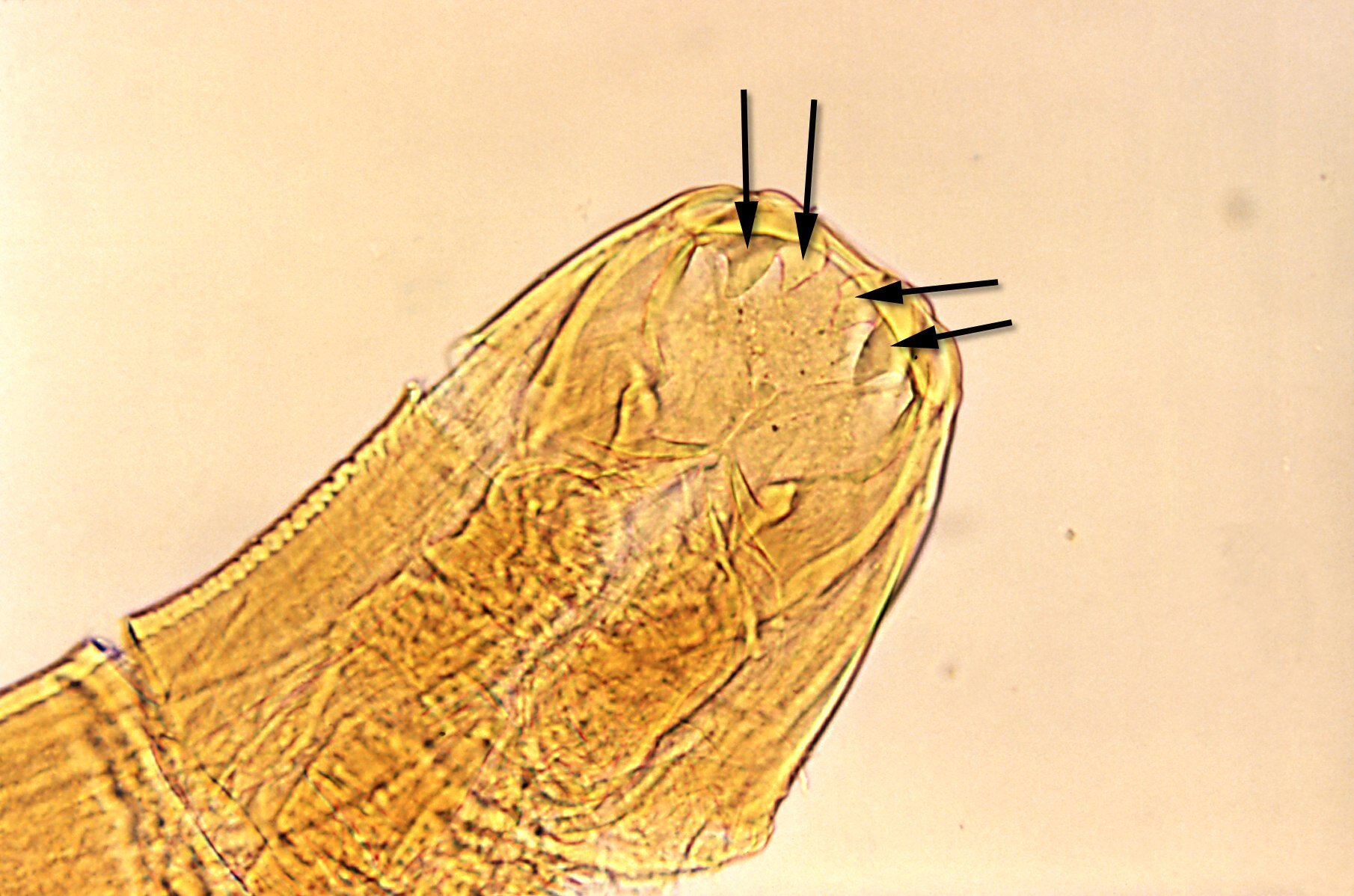
Szereg adaptacji w budowie anatomicznej i fizjologii dotyczy budowy i funkcjonowania układu pokarmowego i rozrodczego oraz przebiegu procesów metabolicznych.
Układ pokarmowy - u pasożytów początkowe odcinki układu pokarmowego zaopatrzone są w struktury umożliwiające pobranie pokarmu.

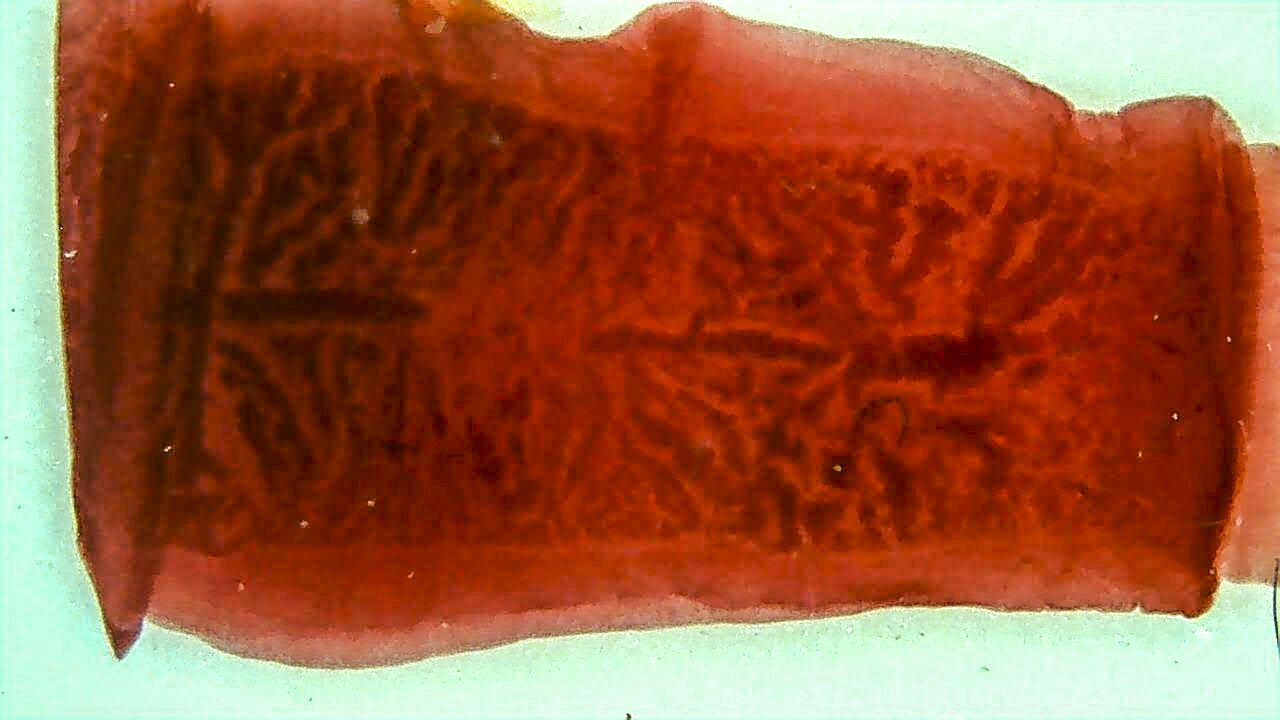
Układ rozrodczy - część pasożytów to obojnakiobojnaki (hermafrodyty). Zdolność do produkcji zarówno plemników, jak i komórek jajowych sprawia, że do rozmnażania płciowego nie jest wymagana obecność drugiego osobnika tego samego gatunku. Zatem pojedynczy osobnik pasożytniczy może przeprowadzić pełen cykl rozwojowy. Obojnactwo wiąże się także ze zwielokrotnieniem liczby narządów rozrodczych, dzięki czemu pasożyt produkuje znaczną liczbę jaj. W ten sposób zwiększa się prawdopodobieństwo przedostania się jaj do organizmu żywiciela pośredniegożywiciela pośredniego, jak również zakażenia większej liczby żywicieli. W konsekwencji wzrastają szanse pasożyta na dokończenie cyklu rozwojowego w organizmie żywiciela ostatecznego.żywiciela ostatecznego.
Część pasożytów ma złożone cykle rozwojowe, w których występuje kilka rodzajów larw, wymagających do dalszego rozwoju różnych gatunków żywicieli pośrednich. Komplikacja cyklu zwiększa rozprzestrzenienie pasożyta w środowisku i tym samym zwiększa jego szanse na osiągnięcie dojrzałości.
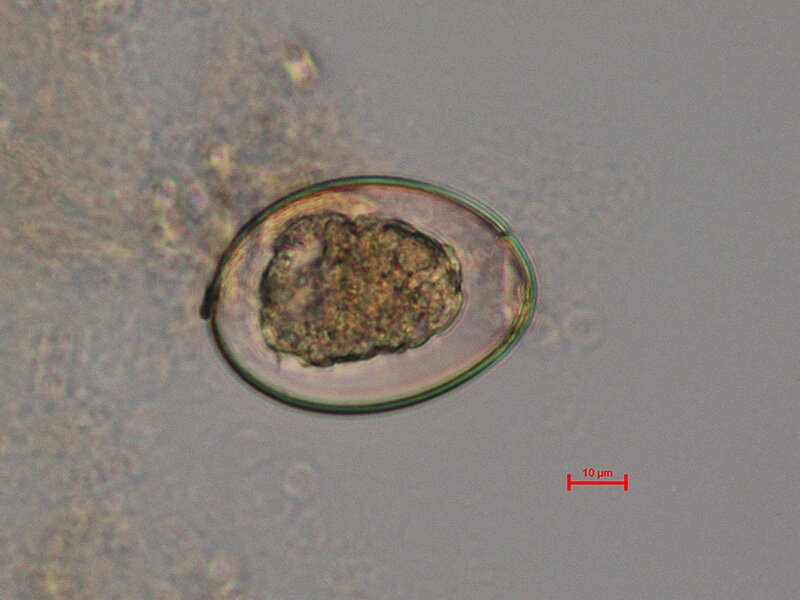
Pasożyty wewnętrzne mają także zdolność oddychania na drodze fermentacji, dzięki czemu mogą pasożytować wewnątrz narządów wewnętrznych, a więc w środowisku ubogim w tlen. Prowadzenie fermentacji jest procesem o niskiej wydajności energetycznej, zatem pokrycie potrzeb energetycznych pasożyta wymaga zużycia znacznych ilości substancji pokarmowych, które pobiera on z ciała żywiciela.
Pasożyty zdolne są do produkcji i wydzielania substancji chemicznych o różnorodnym działaniu. Najczęściej substancje te umożliwiają bezbolesne dla żywiciela pobieranie pokarmu, redukują stan zapalny powstający w miejscu bytowania pasożyta lub zapewniają ochronę przed enzymami trawiennymi gospodarza.

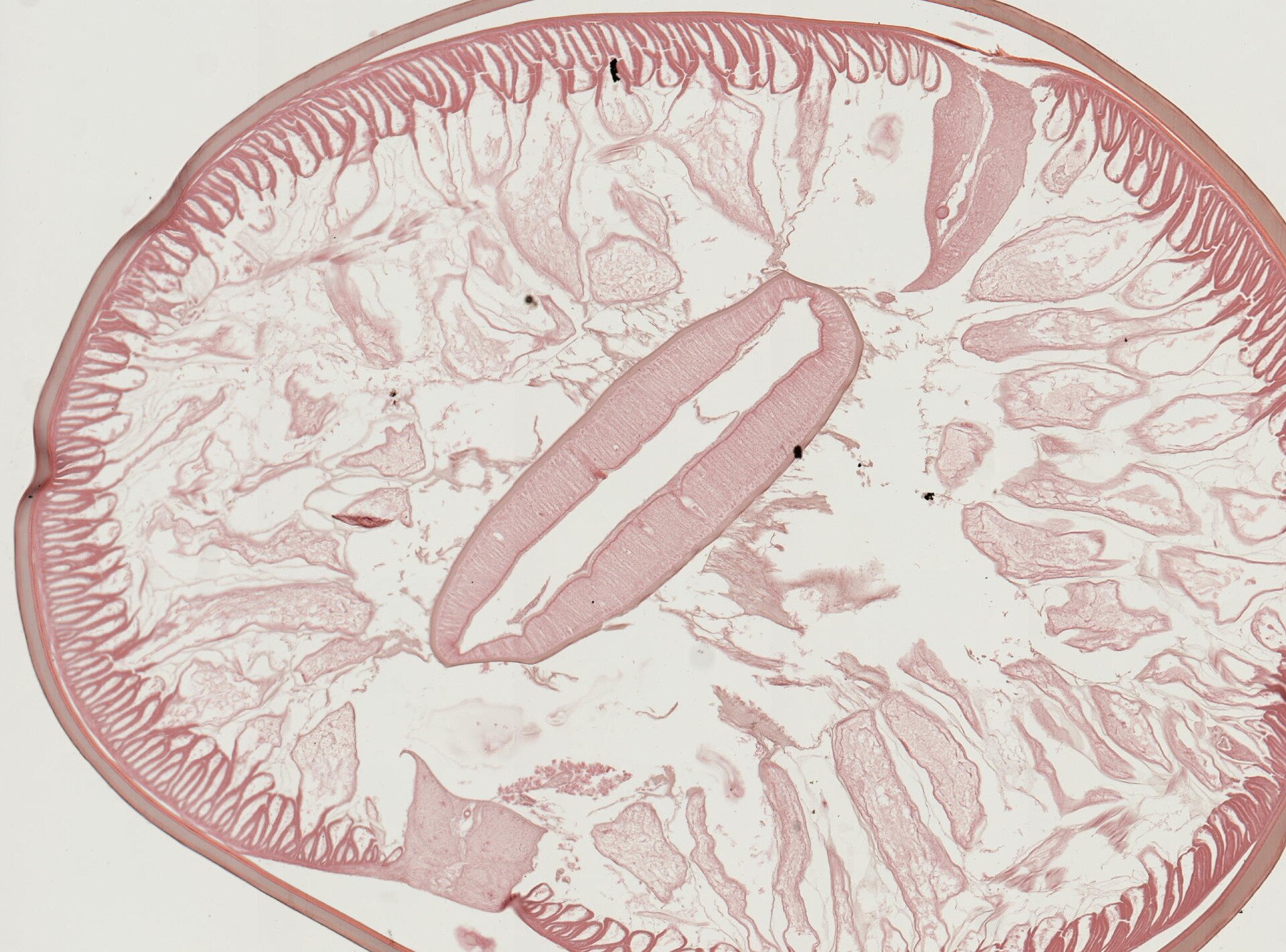
Pasożyt może wykazywać pewne zachowania, które zwiększają prawdopodobieństwo znalezienia żywiciela. Obecność larw pasożyta może wpływać na zmiany w zachowaniu żywiciela pośredniego, dzięki czemu pasożyt zwiększa prawdopodobieństwo przedostania się formy larwalnej do organizmu żywiciela ostatecznego.

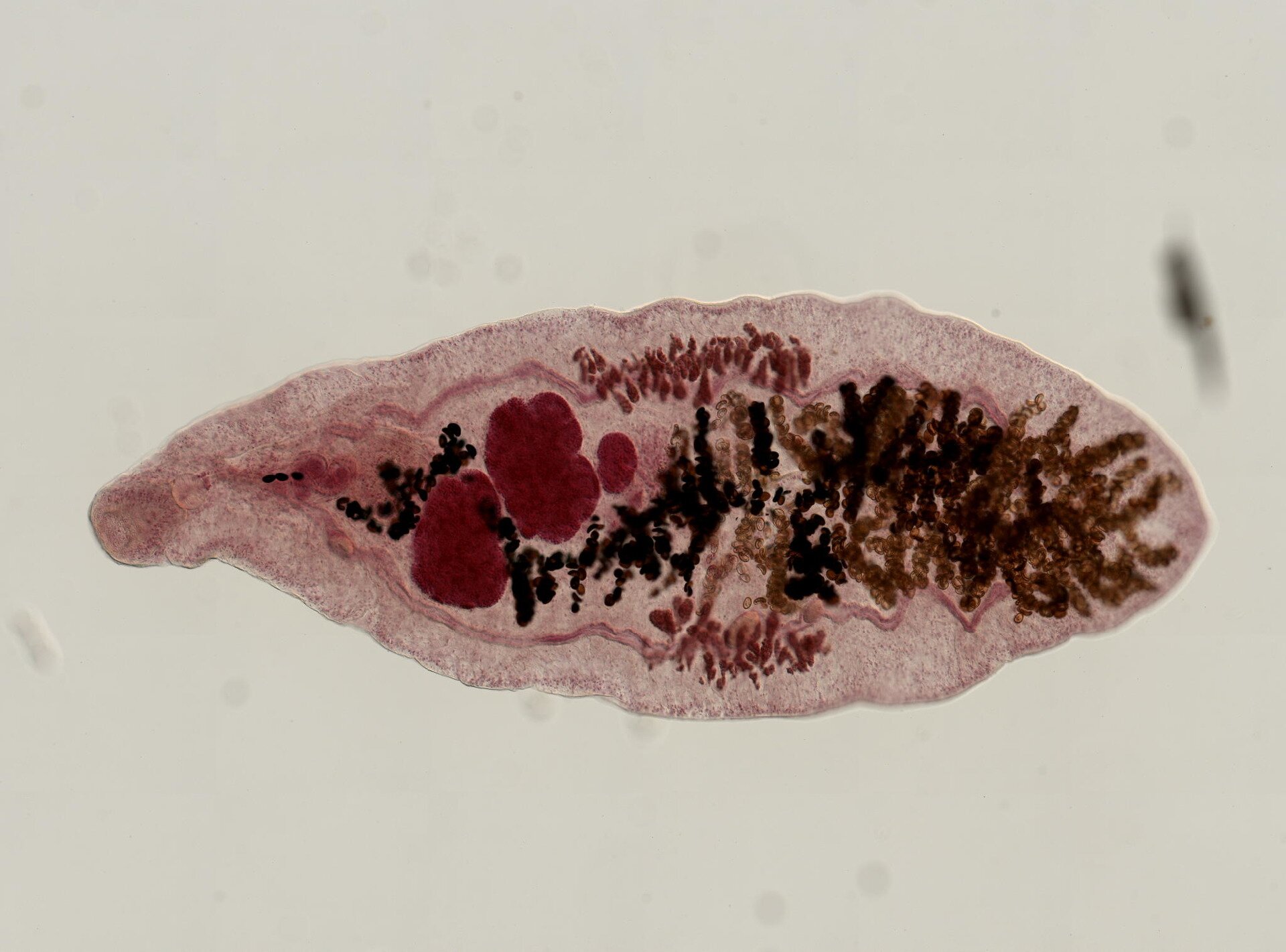
U niektórych ptaków występuje pasożytnictwo lęgowe. Zjawisko to dotyczy m.in. kukułki zwyczajnej (Cuculus canorus), która składa jaja w gniazdach innych gatunków ptaków. Pisklęta kukułek wykluwają się wcześniej niż pisklęta przybranej pary rodzicielskiej. Zazwyczaj usuwają z gniazda właściwe jaja, a niekiedy pisklęta gospodarzy. Wysiłek wykarmienia i odchowania piskląt kukułki podejmują przybrani rodzice.

Przystosowania żywicieli do obrony przed pasożytem
Przystosowania żywicieli do obrony przed pasożytem mają na celu zmniejszenie prawdopodobieństwa infekcji, odstraszenie pasożyta lub jego usunięcie z organizmu. Mechanizmy obronne są możliwe dzięki przystosowaniom: morfologicznym, anatomiczno‑fizjologicznym, chemicznym i behawioralnym.
W budowie morfologicznej żywicieli obecne są struktury, które utrudniają pasożytom wniknięcie do wnętrza ciała danego organizmu. Do przystosowań tych zalicza się m.in.: łuski, pióra, włosy, kutikulę u roślin.


Przystosowania obronne żywicieli w zakresie budowy anatomicznej i fizjologii zmniejszają ryzyko zainfekowania przez pasożyta lub prowadzą do jego zniszczenia i usunięcia. Wyróżnia się nieswoiste i swoiste elementy obrony.
Do nieswoistych elementów obrony należą m.in.:
ciągła i nieuszkodzona powłoka skórna oraz błony śluzowe wyścielające drogi pokarmowe, wydalnicze i rozrodcze;
zdolność do wydzielania potu i łoju;
silnie kwaśny odczyn soku żołądkowego;
obecność lizozymulizozymu w ślinie i łzach;
stan zapalny, podwyższona temperatura ciała oraz zdolność leukocytów do fagocytowania patogenów i wydzielania substancji zabijających pasożyty.
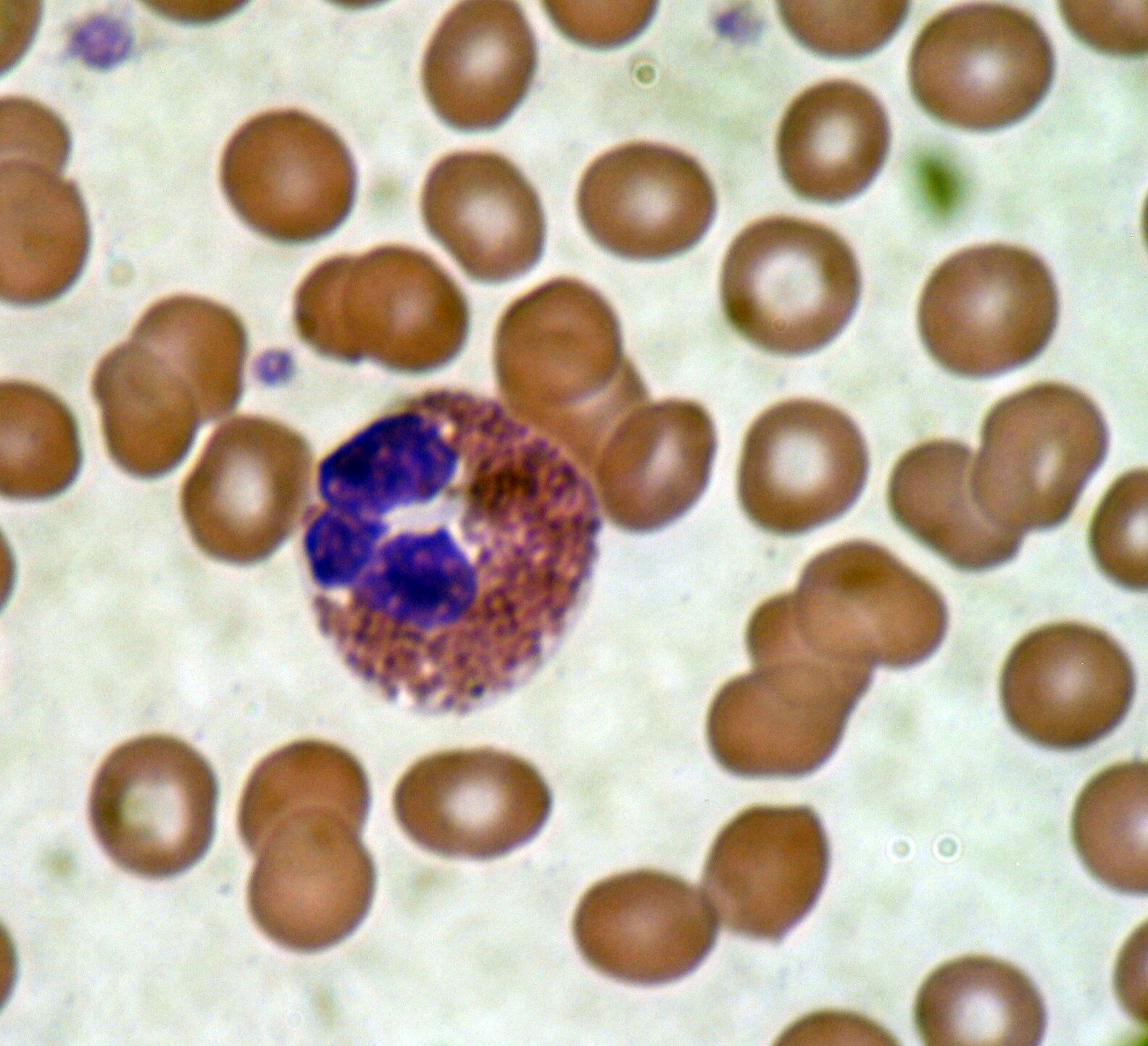
Do swoistych elementów obrony zalicza się zdolność niektórych leukocytów do produkcji przeciwciał neutralizujących patogeny.
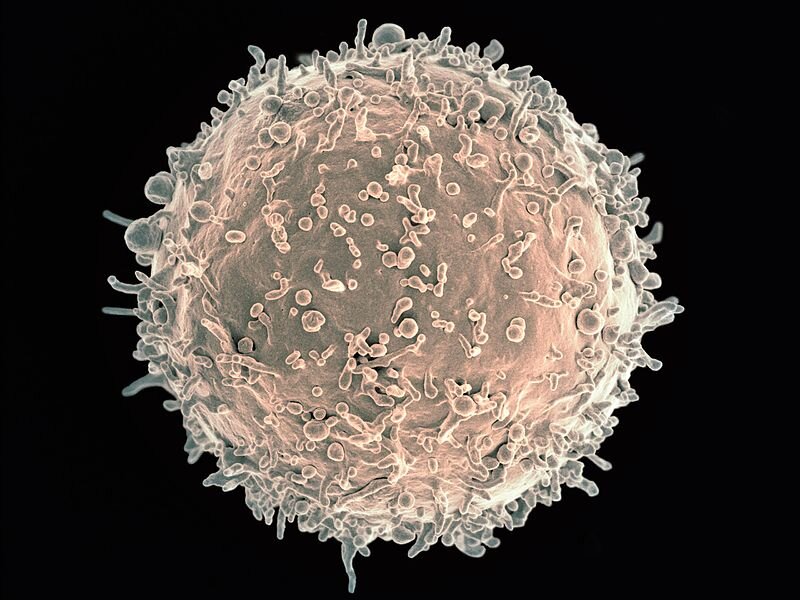
Żywiciele zdolni są do produkcji i wydzielania substancji mających działanie przeciwpasożytnicze lub zniechęcające pasożyty do bytowania w ciele żywiciela.

Obronne przystosowania behawioralne dotyczą różnorodnych zachowań żywicieli związanych z ich funkcjonowaniem. Są to m.in. kąpiele błotne i ocieranie się o przedmioty, mające na celu mechaniczne usunięcie pasożytów zewnętrznych z powierzchni ciała.

Konkurencja
Konkurencja to rodzaj oddziaływania antagonistycznego, w którym dwa organizmy – należące do tego samego lub różnych gatunków – konkurują ze sobą o zasoby środowiska. Konkurencja między roślinami dotyczy dostępu do: światła, wody, soli mineralnych, przestrzeni do życia. Zwierzęta konkurują ze sobą o wodę, pokarm, miejsce i partnera do rozrodu, pozycję w hierarchii stada.
Intensywność konkurencji zależy od stopnia nakładania się nisz ekologicznych konkurujących ze sobą osobników.
Konkurencja może mieć charakter:
eksploatacji, która jest oddziaływaniem pośrednim, w którym osobniki wzajemnie wyczerpują zasoby, o które konkurują;
interferencji, która jest oddziaływaniem bezpośrednim, w którym zwierzęta walczą o terytorium lub samicę, a rośliny stosują walkę chemiczną.
Wyróżnia się:
konkurencję wewnątrzgatunkową;
konkurencję międzygatunkową.
Konkurencja wewnątrzgatunkowa występuje między osobnikami tego samego gatunku, żyjącymi w jednej populacji.
Zależność ta nasila się wraz ze wzrostem liczebności osobników w populacji. Im większe zagęszczenie populacji, tym mniej zasobów środowiska przypada na jednego osobnika.
Konkurencja wewnątrzgatunkowa u roślin prowadzi do:
samoprzerzedzania się - część osobników ginie i zwiększa się dostępność zasobów dla pozostałych osobników.
Konkurencja wewnątrzgatunkowa u roślin i zwierząt prowadzi do:
migracji w poszukiwaniu nowych miejsc nadających się do zasiedlenia, w których zagęszczenie osobników nie jest tak duże. Migrują przede wszystkim osobniki młode, a u roślin – formy specjalnie wytwarzane w celu migracji.


Wynikiem konkurencji wewnątrzgatunkowej u zwierząt jest:
samoregulacja – wraz ze wzrostem liczebności osobników rośnie śmiertelność, a maleje rozrodczość, dlatego zostaje zachowana równowaga w populacji;
terytorializmterytorializm, w wyniku którego teren zajmowany przez populację gatunku podzielony zostaje na mniejsze obszary - terytoria zajmowane przez pojedyncze osobniki lub grupy np. grupy rodzinne. Granice terytorium są oznaczane i bronione przed innymi osobnikami tego samego gatunku;
hierarchiahierarchia społeczna, która sprzyja utrwalaniu grupy i ustala kolejność korzystania z zasobów. Osobniki zajmujące wysokie miejsce w hierarchii mają lepszy dostęp do pożywienia, w związku z czym są większe i silniejsze niż pozostałe.

Gatunki o zbliżonych wymaganiach życiowych i korzystające z tych samych zasobów środowiska mają zbliżone nisze ekologiczne, które mogą się w dużym stopniu pokrywać. Między osobnikami tych gatunków pojawia się rywalizacja o przetrwanie – konkurencja międzygatunkowa.
Konkurencja międzygatunkowa prowadzi do:
ograniczenia niszy ekologicznej jednego z konkurentów;
Dwa gatunki unikające konkurencji bezpośredniej mogą ze sobą współistnieć, jeśli ich nisze nie nakładają się całkowicie. Przykładem są dwa gatunki kormoranów gnieżdżących się na wybrzeżach Wysp Brytyjskich – kormoran czarny (Phalacrocorax carbo) i kormoran czubaty (Phalacrocorax aristotelis) współistnieją na tym samym terenie. Ich nisze ekologiczne są podobne, ale nie identyczne. Każdy z gatunków odżywia się pokarmem z innego obszaru zbiornika wodnego.
konkurencyjnego wyparcia gatunku (zasada Gausegozasada Gausego) z zajmowanego terenu.
Do wyparcia jednej z populacji konkurujących gatunków z niszy ekologicznej nie zawsze dochodzi w wyniku bezpośredniego ataku konkurentów lub wydzielania do środowiska toksycznych substancji. Zwycięstwo odnoszą również osobniki, które skuteczniej wykorzystują zasoby pokarmu i szybciej się rozmnażają.
Przykładem konkurencyjnego wyparcia gatunku wyparcie gatunku wiewiórki rudej (Sciurus vulgaris) przez wiewiórkę szarą (Sciurus carolinensis). Sprowadzenie do Wielkiej Brytanii wiewiórki szarej, która występuje powszechnie w Ameryce Północnej, spowodowało wyparcie rodzimego gatunku wiewiórki rudej. Przyczyną było zajmowanie tych samych nisz ekologicznych.
silnie wpływa na rozmieszczenie konkurujących gatunków w przestrzeni.
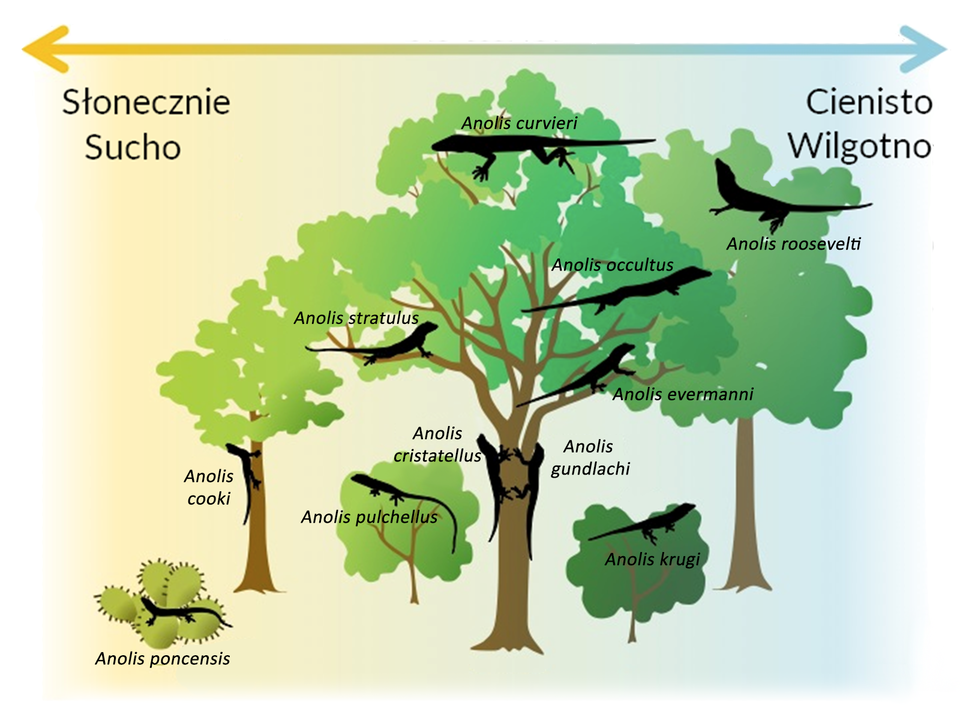
zmian morfologicznych i fizjologicznych w ewoluujących zespołach jednego lub obu gatunków;
doboru rozrywającegodoboru rozrywającego prowadzącego do wyeliminowania osobników o średniej wartości danej cechy i promowaniu osobników o cechach skrajnych, przez co utrzymuje się zróżnicowanie genetyczne i prowadzi do specjacji.
Konkurencja międzygatunkowa jest jednym z podstawowych czynników doboru naturalnego, który jest mechanizmem specjacji i ewolucji biologicznej.
Przeanalizuj wykresy przedstawiające doświadczenie Gausego. Na ich podstawie wykonaj polecenia.
Hipoteza Gausego, inaczej zasada konkurencyjnego wyparcia, zakłada, że dwa gatunki o identycznych wymaganiach, potrzebujące tej samej niszy ekologicznej, nie mogą współistnieć ze sobą i z czasem, w wyniku konkurencji jeden zostaje wyparty. Prawdziwość teorii Gausego została potwierdzona doświadczeniem. Umieszczono w kolbach dwa gatunki pantofelków Paramecium aureliai Paramecium caudatum. Jako źródło pożywienia stosowano kulturę bakteryjną. Pantofelki miały te same wymagania żywieniowe, zatem oba gatunki zostały zmuszone do dzielenia tej samej niszy ekologicznej.
Problemem badawczym eksperymentu Gausego było sprawdzenie jak zachowają się gatunki mające te same potrzeby i żyjące w tym samym środowisku, gdy będą zmuszone konkurować o pokarm.
Kliknij na poszczególne elementy symulacji i obserwuj, jak zmienia się objętość populacji pantofelka Paramecium aurelia w pojedynczej hodowli oraz objętość populacji pantofelka Paramecium aurelia w populacji mieszanej z pantofelkiem Paramecium caudatum.
Zapoznaj się z poniższymi danymi i zawuaż, jak zmienia się objętość populacji pantofelka Paramecium aurelia w pojedynczej hodowli oraz objętość populacji pantofelka Paramecium aurelia w populacji mieszanej z pantofelkiem Paramecium caudatum.
Oś X wyznacza dni prowadzonego doświadczenia, oś Y – objętość populacji.
Dane przedstawiające objętość populacji pojedynczej hodowli pantofelka Paramecium aurelia oraz mieszanej populacji pantofelków Paramecium aurelia i Paramecium caudatum:
Dzień 0. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 0. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 0.
Dzień 2. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 12. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 12.
Dzień 4. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 26. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 25.
Dzień 6. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 44. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 40.
Dzień 8. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 110. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 75.
Dzień 10. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 165. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 100.
Dzień 12. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 210. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 125.
Dzień 14. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 215. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 127.
Dzień 16. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 210. OObjętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 120.
Dzień 18. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 205. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 114.
Dzień 20. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 200. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 110.
Dzień 22. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 198. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 105.
Dzień 24. Objętość populacji Paramecium aurelia hodowanej oddzielnie: 195. Objętość populacji Paramecium aurelia w hodowli mieszanej z Paramecium caudatum: 100.
Oś X wyznacza dni prowadzonego doświadczenia, oś Y – objętość populacji.
Na podstawie: William P. Cunningham, Mary Cunningham, Environmental Science: A Global Concern, McGraw‑Hill Science/Engineering/Math 2012 (textbook)..
Dane przedstawiające objętość populacji pojedynczej hodowli pantofelka Paramecium caudatum oraz mieszanej populacji pantofelków Paramecium aurelia i Paramecium caudatum:
Dzień 0. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 0. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 0.
Dzień 2. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 12. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 12.
Dzień 4. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 25. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 25.
Dzień 6. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 60. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 80.
Dzień 8. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 150. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 125.
Dzień 10. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 190. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 110.
Dzień 12. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 200. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 80.
Dzień 14. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 200. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 65.
Dzień 16. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 200. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 60.
Dzień 18. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 200. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 50.
Dzień 20. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 200. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 40.
Dzień 22. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 200. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 25.
Dzień 24. Objętość populacji Paramecium caudatum hodowanej oddzielnie: 200. Objętość populacji Paramecium caudatum w hodowli mieszanej z Paramecium aurelia: 20.
Zasada Gausego
Dwa gatunki mogą przetrwać wspólnie w danym miejscu tylko wtedy, gdy zajmują różne nisze ekologiczne.
Allelopatia
Allelopatia to rodzaj oddziaływania antagonistycznego, w którym jeden organizm wydziela do otoczenia substancje allelopatyczne (allelopatiny), ograniczające lub hamujące wzrost i rozwój drugiego organizmu. Organizm wydzielający allelopatiny odnosi korzyści, ponieważ poprzez ograniczenie występowania drugiego uzyskuje dostęp do większych zasobów środowiska. Organizm wrażliwy na allelopatiny ponosi stratę, ponieważ obecność związków chemicznych w środowisku ogranicza jego występowanie.
Substancje allelopatyczne to związki chemiczne będące metabolitami wtórnymi produkowanymi przez mikroorganizmy i rośliny wyższe. Pod względem chemicznym allelopatiny są różnego rodzaju związkami, takimi jak: cyjanowodór, amoniak, kwasy organiczne, antybiotyki i inne.
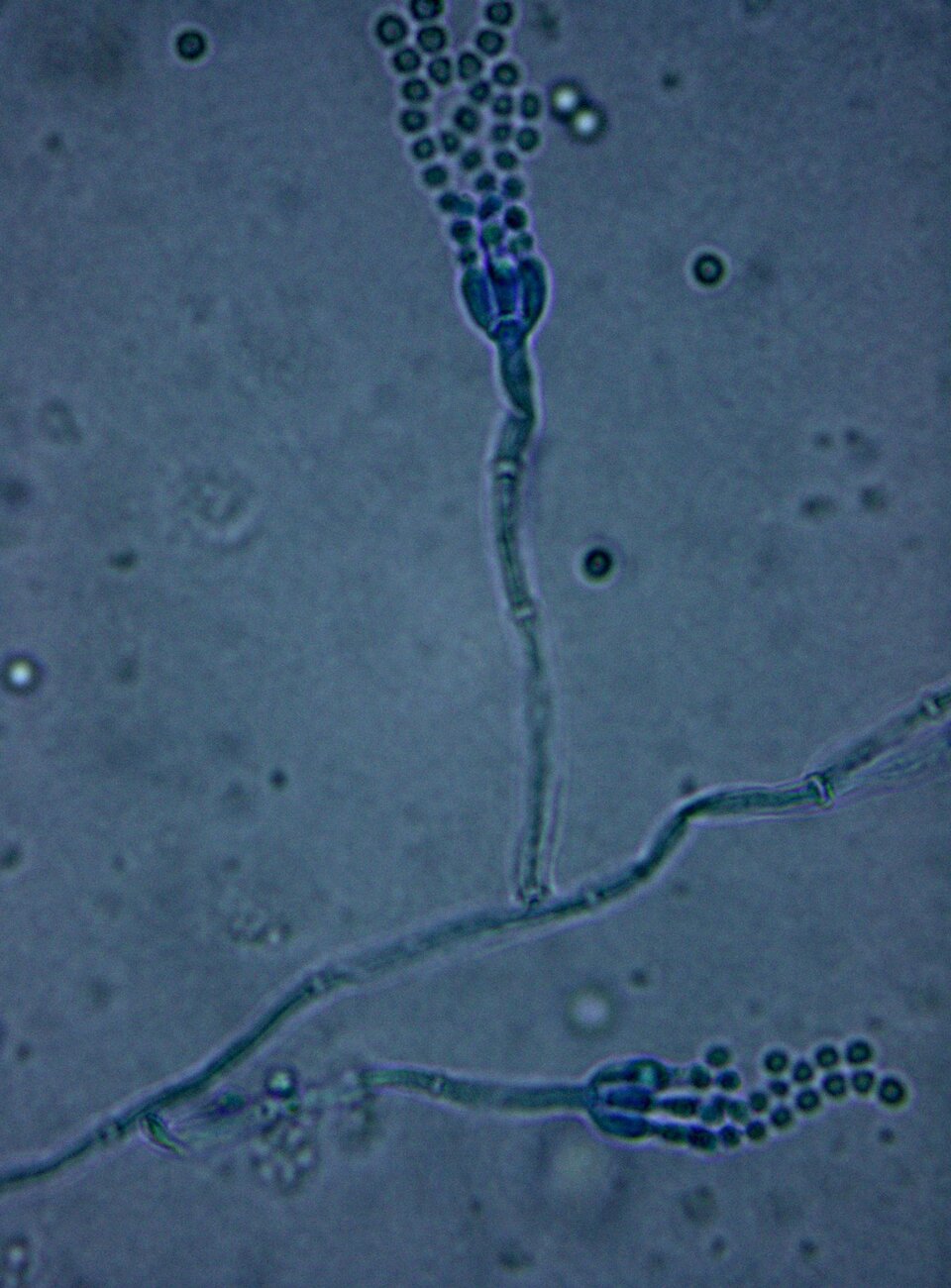
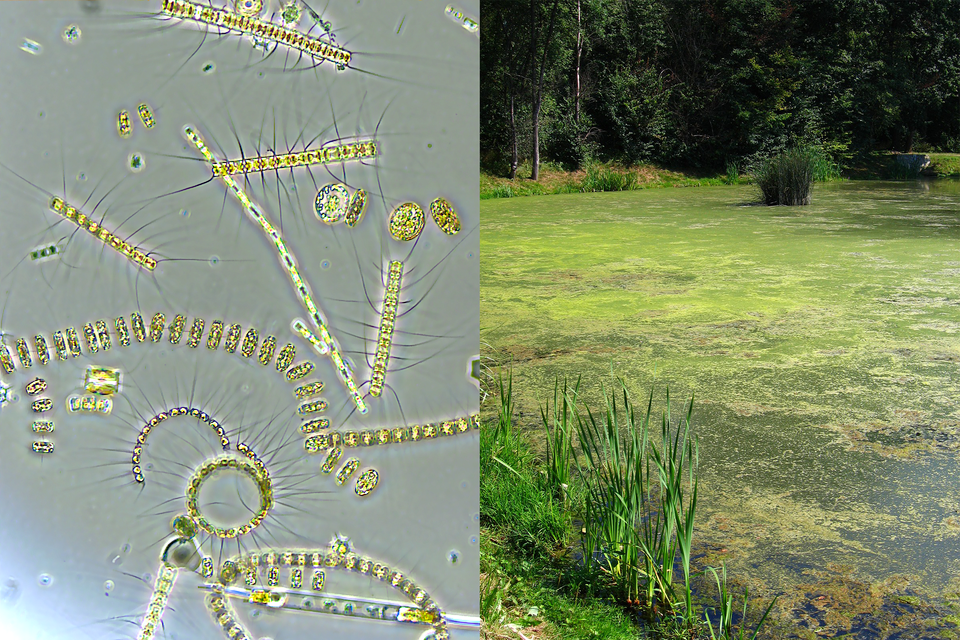
Amensalizm
Amensalizm to rodzaj jednostronnego oddziaływania antagonistycznego, w którym jeden organizm ponosi straty, a drugi w tej zależności ani nie odnosi korzyści, ani nie ponosi strat.
Przykładem może być sytuacja, gdy jeden organizm wydziela do środowiska substancje hamujące wzrost innych organizmów (np. antybiotyki produkowane przez niektóre bakterie), nie odnosząc z tego bezpośredniej korzyści.
Podsumowanie
Zależności antagonistyczne – to oddziaływania między organizmami, w których co najmniej jedna strona ponosi straty. Należą do nich: konkurencja, drapieżnictwo, roślinożerność, pasożytnictwo, amensalizm i allelopatia. Regulują liczebność populacji, wpływają na strukturę ekosystemu i przebieg doboru naturalnego.
Zależności antagonistyczne pełnią kluczową rolę w utrzymaniu równowagi biologicznej, regulują liczebność populacji i przyczyniają się do powstawania przystosowań ewolucyjnych organizmów.
Konkurencja
Zachodzi, gdy organizmy korzystają z tych samych, ograniczonych zasobów (pokarm, woda, światło, przestrzeń, partner do rozrodu).
Wewnątrzgatunkowa – między osobnikami tego samego gatunku; prowadzi do samoregulacji populacji (spadek rozrodczości, wzrost śmiertelności), migracji, terytorializmu, ustalania hierarchii społecznych; u roślin powoduje samoprzerzedzanie.
Międzygatunkowa – między gatunkami o podobnej niszy ekologicznej; może prowadzić do ograniczenia niszy lub wyparcia jednego z gatunków (zasada konkurencyjnego wypierania Gausego).
Drapieżnictwo
Zależność, w której drapieżnik zabija i zjada ofiarę.
Ma duże znaczenie regulacyjne – wpływa na liczebność populacji ofiar i selekcję osobników słabszych.
W układzie drapieżca–ofiara liczebność populacji zmienia się cyklicznie: wzrost liczby ofiar powoduje wzrost liczby drapieżników, a spadek ofiar – spadek drapieżników; maksimum liczebności drapieżników występuje z opóźnieniem względem maksimum ofiar (ujemne sprzężenie zwrotne).
Roślinożerność
Odmiana drapieżnictwa, w której organizm (roślinożerca) zjada całą roślinę lub jej część.
Zwykle nie prowadzi do natychmiastowej śmierci rośliny, ale ogranicza jej wzrost i zdolność do rozmnażania.
Ma znaczenie w regulacji biomasy roślin oraz przepływie energii w ekosystemie.
Pasożytnictwo
Zależność, w której pasożyt żyje kosztem żywiciela, czerpiąc z niego pokarm i osłabiając go, ale zazwyczaj nie doprowadzając szybko do jego śmierci.
Pasożyty mogą być zewnętrzne (ektopasożyty) lub wewnętrzne (endopasożyty).
Pasożyty często wykazują dużą płodność, obecność narządów czepnych i złożone cykle rozwojowe.
Amensalizm
Zależność, w której jeden organizm ponosi straty, a drugi nie odnosi ani korzyści, ani strat.
Allelopatia
Polega na wydzielaniu do środowiska substancji chemicznych hamujących kiełkowanie lub wzrost innych roślin.
Adaptacje do zdobywania pokarmu
Drapieżniki – kły, pazury, jad, silne mięśnie, rozwinięte zmysły, strategie polowania (stadne, z zaskoczenia).
Pasożyty – narządy czepne, redukcja niektórych narządów, duża liczba potomstwa.
Roślinożercy – przystosowane uzębienie, wydłużony przewód pokarmowy, symbioza z mikroorganizmami trawiącymi celulozę.
Mechanizmy obronne ofiar, żywicieli i roślin
Morfologiczne – kolce, pancerze, ubarwienie ochronne.
Anatomiczno‑fizjologiczne – sprawne zmysły, reakcje odpornościowe.
Chemiczne – toksyny, jad, substancje odstraszające.
Behawioralne – ucieczka, życie w grupie, autotomia, zachowania higieniczne.
Ćwiczenia utrwalające
Na wykresie przedstawiono zmiany liczebności populacji drapieżcy i ofiary.
Spośród wymienionych cech wybierz i zaznacz te, które charakteryzują wyłącznie drapieżnika. Możliwe odpowiedzi: 1. zabija ofiarę, 2. osłabia ofiarę, 3. najczęściej większy od swoje ofiary, 4. zawsze mniejszy od swojej ofiary, 5. stosuje kamuflaż
„Akacje bez problemu tolerują stratę do 10% swoich liści. Gdy jednak te straty sięgają 40–50%, akacje nagle zaczynają produkować ogromne ilości tanin i etylenu. Badacze sawanny stwierdzili, że ten gaz niesie informację o zagrożeniu do drzew oddalonych nawet o 100 m. Co ciekawe, w naturalnych warunkach roślinożercy sawanny wiedzą, ile mogą bezpiecznie skubnąć z jednego drzewa. Do zatruć zwierzyny dochodzi na zamkniętych farmach, gdzie zbyt duże zagęszczenie zwierząt zmusza je do ogołocenia nielicznych drzew”.
Indeks górny Źródło: M. Mazurek, Drzewa mają niesamowity system obrony, www.plus.gazetakrakowska.pl. Materiał wykorzystany na podstawie art. 29 ustawy o prawie autorskim i prawach pokrewnych (prawo cytatu). Indeks górny koniecŹródło: M. Mazurek, Drzewa mają niesamowity system obrony, www.plus.gazetakrakowska.pl. Materiał wykorzystany na podstawie art. 29 ustawy o prawie autorskim i prawach pokrewnych (prawo cytatu).
Na podstawie powyższego tekstu zaznacz wszystkie prawidłowe stwierdzenia.
Wróć do polecenia na stronie „Na dobry początek” i dopisz brakujące definicje. Pamiętaj, żeby nie kopiować słownika, ale wyjaśnić każde słowo kluczowe w miarę możliwości swoimi słowami.