Genetyka mendlowska
Chromosomowa teoria dziedziczenia i geny sprzężone
Przedstawisz główne założenia chromosomowej teorii dziedziczenia Thomasa Morgana.
Wykażesz różnicę między genami sprzężonymi i genami niesprzężonymi.
Omówisz pojęcie organizmu modelowego na przykładzie muszki owocowej.
Obliczysz odległość między genami oraz określisz ich ułożenie na chromosomie.
Niepozorna muszka owocowa (Drosophila melanogaster) stała się obiektem epokowych badań, nagrodzonych w 1933 r. Nagrodą Nobla. Na początku XX w. Thomas Morganpodczas eksperymentów prowadzonych na muszkach owocowych zaobserwował mutację powiązaną z płcią w genie wpływającym na kolor oczu: część okazów muszek miała białe oczy, zamiast charakterystycznych dla tego gatunku oczu czerwonych. Analiza dziedziczenia tej cechy, według innego wzoru u samców i u samic muszek, dała ostateczny dowód na to, że geny znajdują się w chromosomach.
Przypomnij sobie budową chromosomu
Założenia chromosomowej teorii dziedziczenia
Thomas Morgan i jego współpracownicy badając muszkę owocową wykazali, że:
chromosomy są zbiorami ułożonych liniowo genów, które nie zachodzą na siebie;
gen zajmuje w chromosomie ściśle określone miejsce, tzw. locus (l. mn.: loci), a allele jednego genu – warunkującego określoną cechę – zajmują to samo miejsce w chromosomach homologicznych;
geny znajdujące się w tym samym chromosomie nazywane są genami sprzężonymi i są dziedziczone razem;
geny sprzężone mogą ulec rozdzieleniu na skutek zachodzącego podczas mejozy procesu zwanego crossing‑over, polegającego na wymianie chromatyd między chromosomami homologicznymi.
Organizm modelowy to gatunek, który służy naukowcom do badania i opisywania uniwersalnych mechanizmów biologicznych, takich jak dziedziczenie cech czy podłoże genetyczne chorób. W genetyce zwierząt funkcję tę pełnią najczęściej muszka owocowa (Drosophila melanogaster) oraz nicień Caenorhabditis elegans. Z kolei najważniejszym modelem w badaniach nad genetyką roślin jest rzodkiewnik pospolity (Arabidopsis thaliana)



Geny sprzężone i niesprzężone
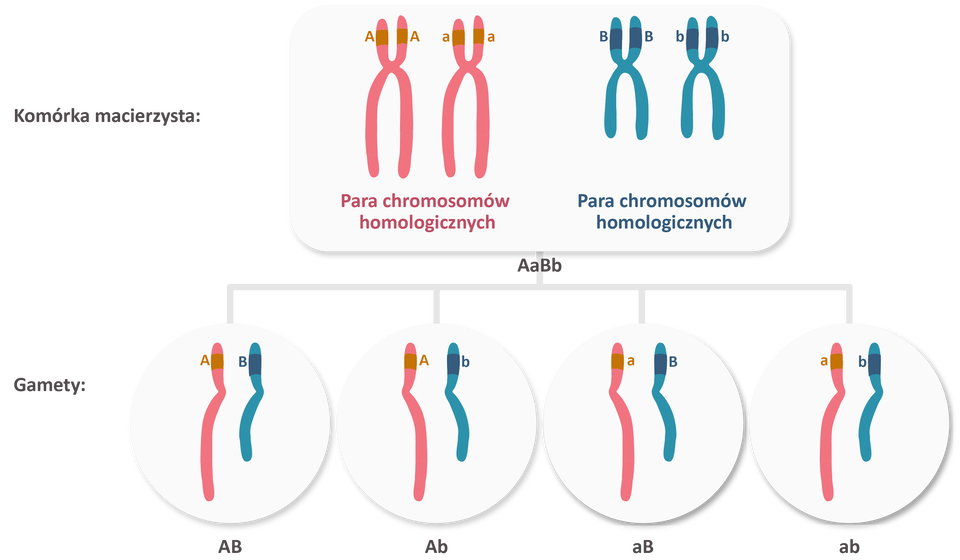
Większość chromosomów zawiera setki lub nawet tysiące genów. Geny znajdujące się na jednym chromosomie nazywane są genami sprzężonymi. Geny niesprzężone natomiast znajdują się na różnych chromosomach.

Geny niesprzężone dziedziczą się niezależnie od siebie, zgodnie z II prawem Mendla. Dzieje się tak, ponieważ znajdują się na chromosomach niehomologicznych, które rozchodzą się do gamet w sposób losowy.
Geny sprzężone, natomiast zazwyczaj przechodzą do gamety razem (wraz z chromosomem, na którym się znajdują) , dlatego ich dziedziczenie jest niezgodne z II prawem Mendla.
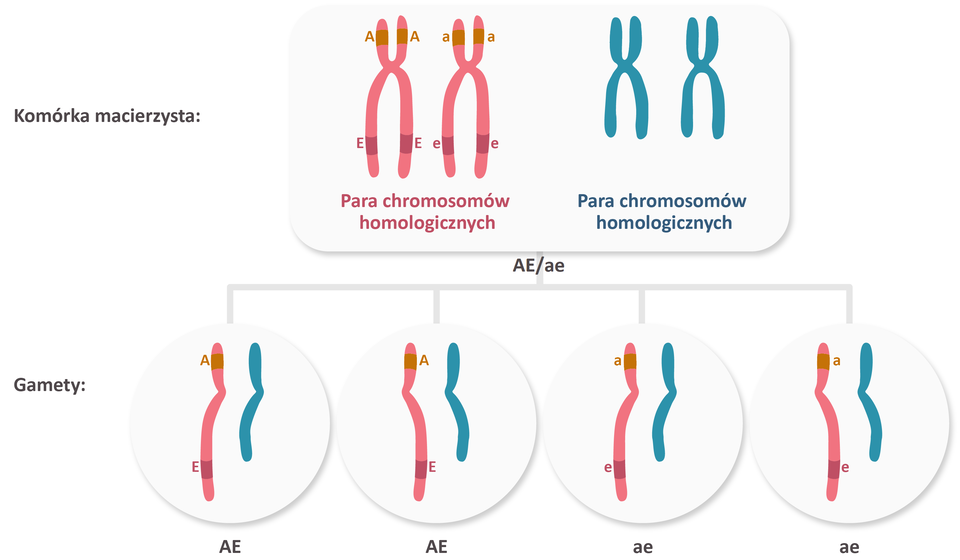
Sprzężenie genów oznacza się zazwyczaj za pomocą kreski ułamkowej, gdzie allele zlokalizowane na jednym z chromosomów homologicznych umieszcza się nad kreską, a allele zlokalizowane na drugim chromosomie homologicznym – pod kreską. Np. zapis AB/ab oznacza, że jednym chromosomie znajdują się allele A i B, a na drugim - a i b. Ponieważ geny sprzężone dziedziczą się razem, allele znad kreski ułamkowej przedostają się jednej gamety, a allele spod kreski - do drugiej.
Sprzężenie genów oznacza się zazwyczaj za pomocą kreski ułamkowej, gdzie allele zlokalizowane na jednym z chromosomów homologicznych umieszcza się nad kreską, a allele zlokalizowane na drugim chromosomie homologicznym – pod kreską. Np. zapis AB/ab (duże A duże Be kreska małe a małe b) oznacza, że jednym chromosomie znajdują się allele A (duże A) i B (duże Be), a na drugim - a (małe a) i b (małe be). Ponieważ geny sprzężone dziedziczą się razem, allele znad kreski ułamkowej przedostają się jednej gamety, a allele spod kreski - do drugiej.
Obliczanie prawdopodobieństwa dziedziczenia cech sprzężonych
Analiza fenotypów potomstwa pozwala ustalić, czy geny dziedziczą się zgodnie z II prawem Mendla (są niesprzężone), czy są ze sobą sprzężone. W przypadku genów niesprzężonych (leżących na różnych chromosomach), skrzyżowanie podwójnych heterozygot daje w pokoleniu FIndeks dolny 22 cztery klasy fenotypowe w stosunku 9:3:3:1.
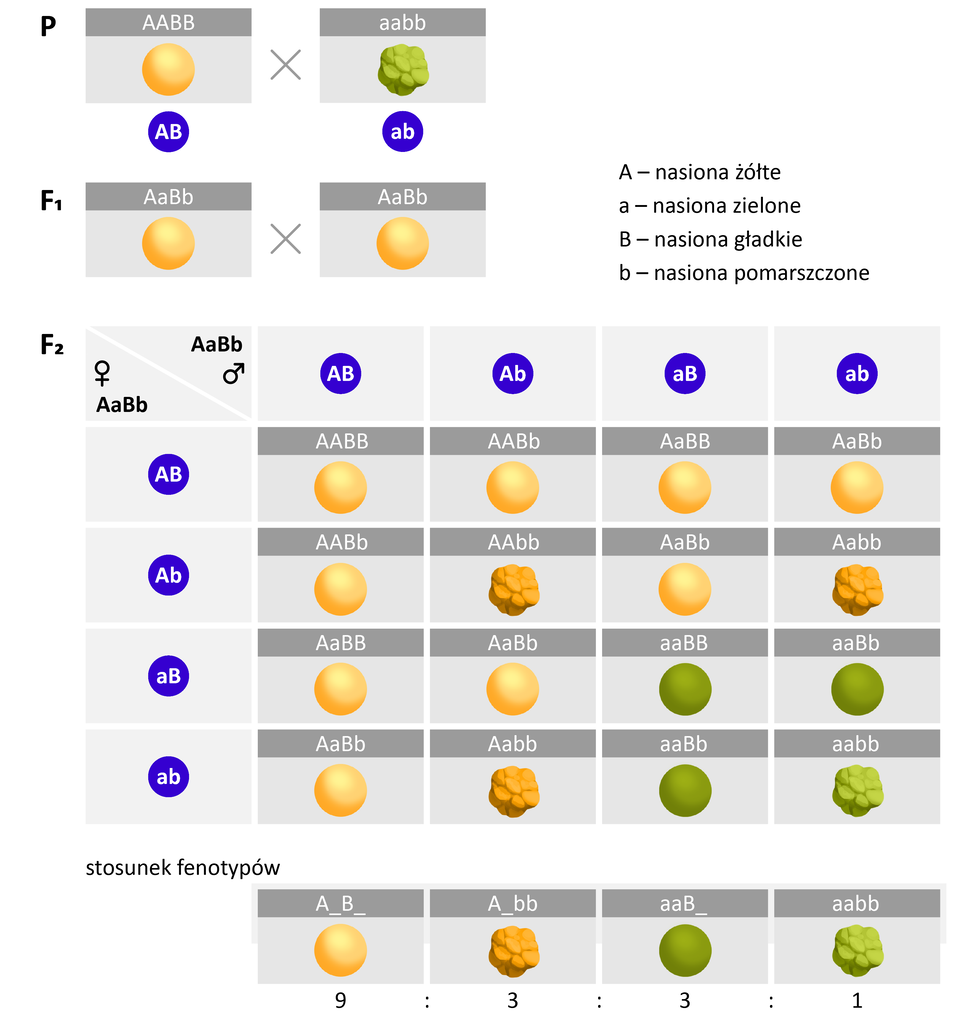
Gdyby jednak te same geny były sprzężone (leżały na tej samej parze chromosomów), w pokoleniu FIndeks dolny 22 pojawiłyby się tylko dwie klasy fenotypowe w stosunku 3:1. Oznacza to, że dwie różne cechy sprzężone dziedziczą się tak, jakby były jedną cechą.
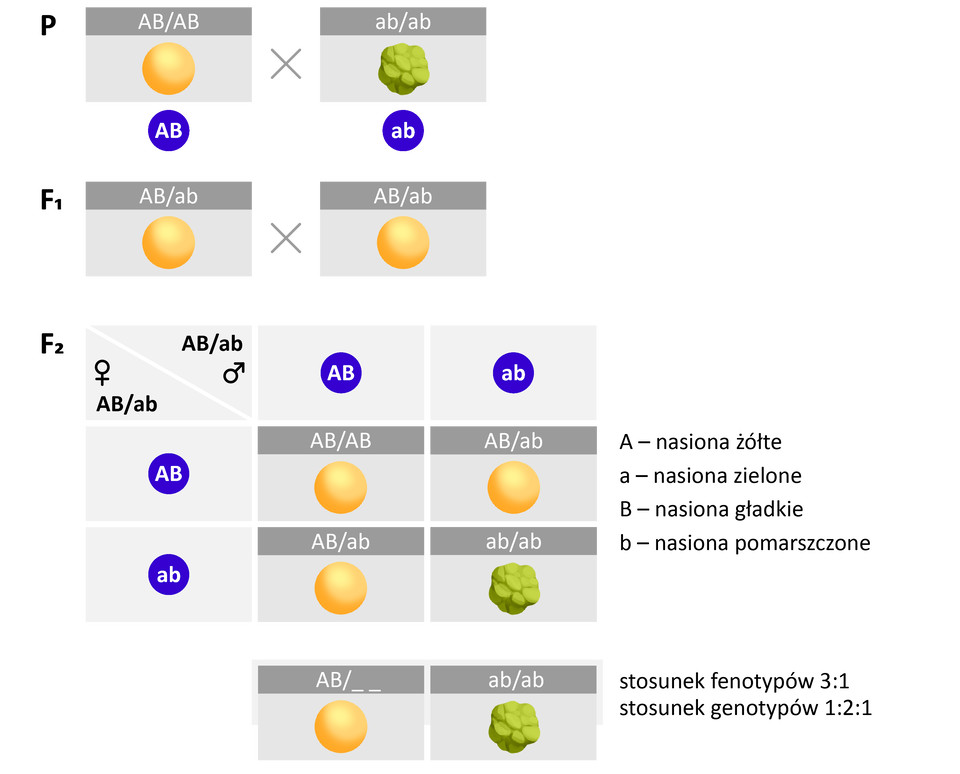
W przypadku sprzężenia cech w pokoleniu potomnym podwójnych heterozygot obserwowane są dwa rodzaje fenotypów osobników potomnych w stosunku 3:1.
Skrzyżowano dwie podwójnie heterozygotyczne muszki owocowe (genotyp DdBb) o normalnych odnóżach i brązowym ciele. W tabeli przedstawiono uzyskany rozkład fenotypów wśród muszek z pokolenia FIndeks dolny 11.
Skrzyżowano dwie podwójnie heterozygotyczne muszki owocowe (genotyp DdBb - duże De małe de duże Be małe be) o normalnych odnóżach i brązowym ciele. W tabeli przedstawiono uzyskany rozkład fenotypów wśród muszek z pokolenia efIndeks dolny 11.
D – normalne odnóża
d – krótkie odnóża
B – brązowe ciało
b – czarne ciało
D (duże De) – normalne odnóża
d (małe de) – krótkie odnóża
B (duże Be) – brązowe ciało
b (małe be) – czarne ciało
Fenotyp potomstwa | Liczba osobników |
|---|---|
Normalne odnóża, brązowe ciało | 295 |
Krótkie odnóża, brązowe ciało | 9 |
Normalne odnóża, czarne ciało | 12 |
Krótkie odnóża, czarne ciało | 98 |
Określ, czy analizowane cechy są ze sobą sprzężone. Odpowiedź uzasadnij.
Rozdzielenie genów sprzężonych
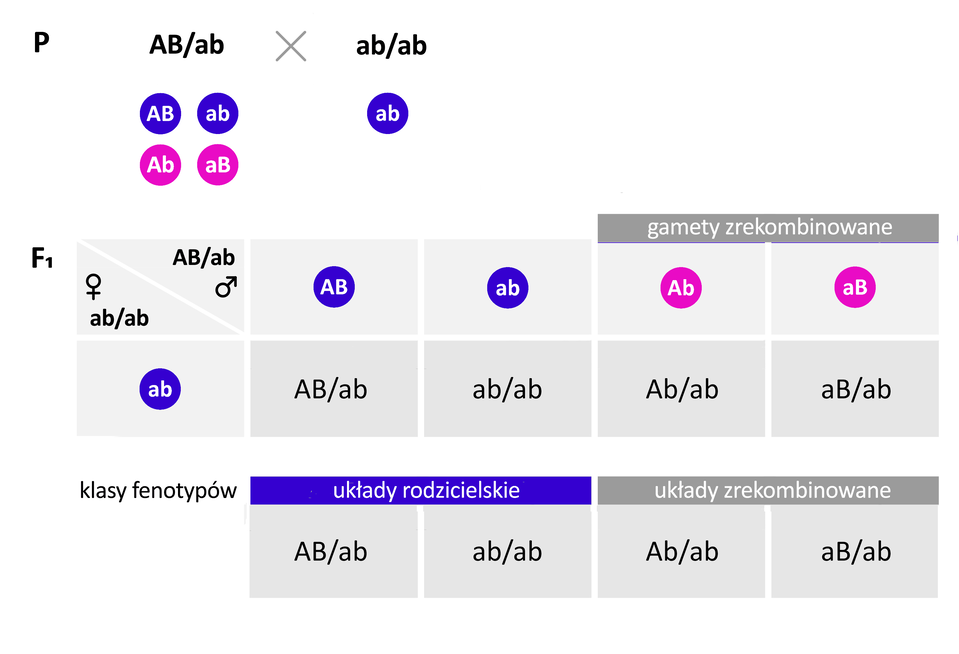
Sprzężenie genów nie jest całkowite. Rozdzielenie genów sprzężonych może nastąpić na skutek crossing‑over, który polega na wymianie chromatyd między chromosomami homologicznymi podczas mejozy. Dzięki temu procesowi allele tego samego genu mogą „zamienić się miejscami” na chromosomach homologicznych. W rezultacie crossing‑over umożliwia wytworzenie nowych układów alleli. Prowadzi do powstania czterech, zamiast dwóch, typów gamet: dwóch takich jak rodzicielskie i dwóch o nowym układzie alleli.
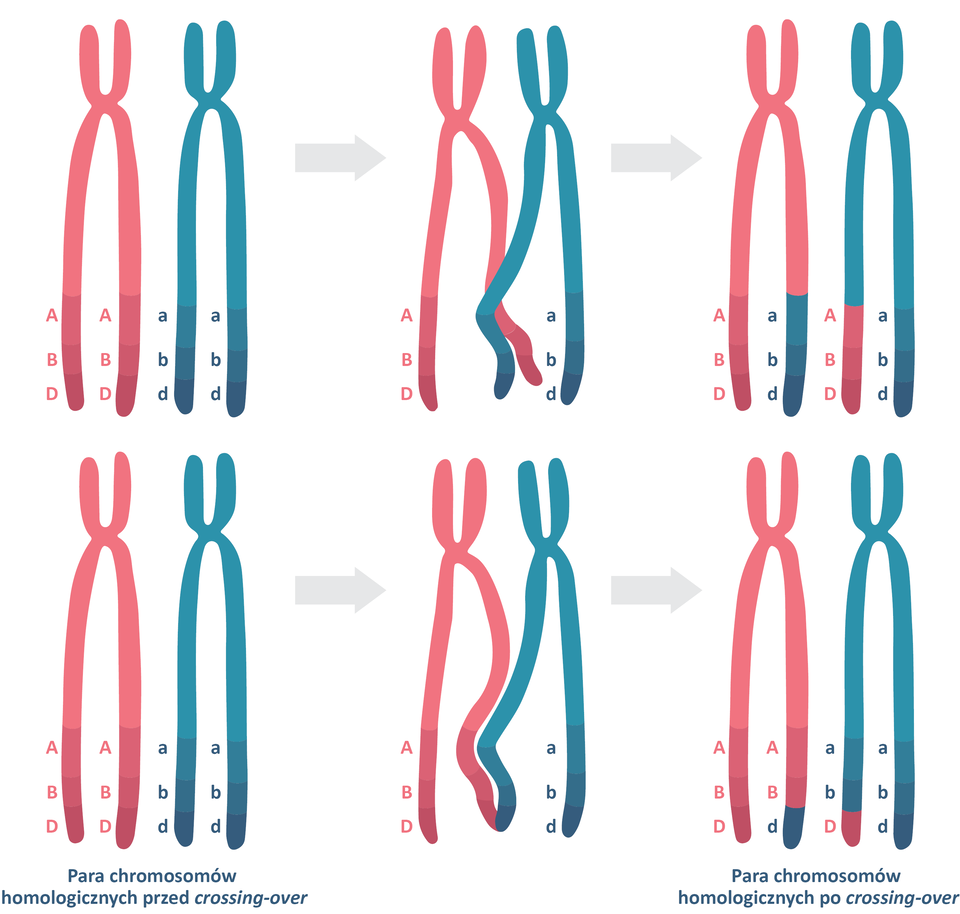
Prawdopodobieństwo wystąpienia crossing‑over jest wprost proporcjonalne do odległości dzielącej geny na chromosomie. Gdy geny leżą blisko siebie, zwykle trafiają do gamet razem, ponieważ szansa na ich rozdzielenie jest niewielka.
Przeanalizuj animację „Obliczanie prawdopodobieństwa dziedziczenia cech sprzężonych” i przedstawione tam zadanie. Następnie spróbuj wykonać polecenie i ćwiczenie.

Film dostępny pod adresem /preview/resource/RL9NMOVA4RE18
Film prezentujący obliczanie prawdopodobieństwa dziedziczenia cech sprzężonych.

Zapoznaj się z animacją „Wykrywanie sprzężenia za pomocą krzyżówki dwugenowej”. Później wykonaj polecenie.

Film dostępny pod adresem /preview/resource/RTWakoMOFnU75
Nagranie filmowe pod tytułem Wykrywanie sprzężenia za pomocą testowej krzyżówki dwugenowej.
Obliczanie częstości zachodzenia crossing‑over
Na podstawie liczby rekombinantówrekombinantów można określić częstość zachodzenia crossing‑over pomiędzy dwoma genami oraz odległość między nimi na chromosomie. Obliczeń tych dokonuje się na podstawie wyników krzyżówki testowej.
Przeprowadza się je następująco:
Poniżej przedstawiono przykład obliczeń częstości crossing‑over pomiędzy genami b i vg u muszki owocowej. Allele warunkujące wystąpienie cech typu dzikiego zapisuje się ze znakiem „+”, natomiast allele odpowiadające za wystąpienie mutacji - bez plusa. Zapis bIndeks górny ++oznacza więc allel warunkujący szarą barwę ciała (fenotyp dziki), natomiast allel b (od angielskiego black) – allel warunkujący wystąpienie mutacji objawiającej się czarną barwą ciała. Allel warunkujący skrzydła normalnej długości zapisuje się jako vgIndeks górny ++, a allel objawiający się zredukowanymi skrzydłami vg (od angielskiego vestigial).
Poniżej przedstawiono przykład obliczeń częstości crossing‑over pomiędzy genami b (małe be) i vg (małe fał małe gie) u muszki owocowej. Allele warunkujące wystąpienie cech typu dzikiego zapisuje się ze znakiem „+”, (plus) natomiast allele odpowiadające za wystąpienie mutacji - bez plusa. Zapis bIndeks górny ++(małe be plus) znacza więc allel warunkujący szarą barwę ciała (fenotyp dziki), natomiast allel b (małe be); od angielskiego black) – allel warunkujący wystąpienie mutacji objawiającej się czarną barwą ciała. Allel warunkujący skrzydła normalnej długości zapisuje się jako vgIndeks górny ++ (małe fał małe gie plus) a allel objawiający się zredukowanymi skrzydłami vg (małe fał małe gie; od angielskiego vestigial).
Obliczenie częstości crossing‑over wymaga wielu analiz statystycznych na dużej liczbie badanych osobników. Wartość tę podaje się w procentach. Na przykład crossing‑over wynoszące 10% oznacza, że u 10% potomstwa nastąpi rozdzielenie sprzężonych ze sobą alleli badanych genów.
Zapoznaj się z filmem „Chromosomowa teoria dziedziczności Morgana”, a następnie wykonaj polecenia.

Film dostępny pod adresem /preview/resource/R1LL4SH9U7MN3
Film nawiązujący do treści materiału - dotyczy chromosomowej teorii dziedziczności Morgana na podstawie obserwacji muszki owocowej.
Mapowanie genów
Znając częstość zachodzenia crossing‑over między dwoma konkretnymi genami, można obliczyć zarówno odległość między tymi genami na chromosomie, jak i określić kolejność ich położenia.
Amerykański biolog Thomas Morgan, który opracował chromosomową teorię dziedziczenia cech, został upamiętniony w nazwie jednostki odległości między genami: 1 centymorgan (cM), inaczej 1 jednostka mapowa (j.m.), odpowiada 1% częstości crossing‑over.
Graficzne przedstawienie umiejscowienia poszczególnych genów na chromosomie nazywa się mapą genetyczną, a obliczenia – mapowaniem chromosomu lub mapowaniem genów.

Przykład obliczania odległości między poszczególnymi genami na chromosomie oraz ustalania ich położenia na chromosomie
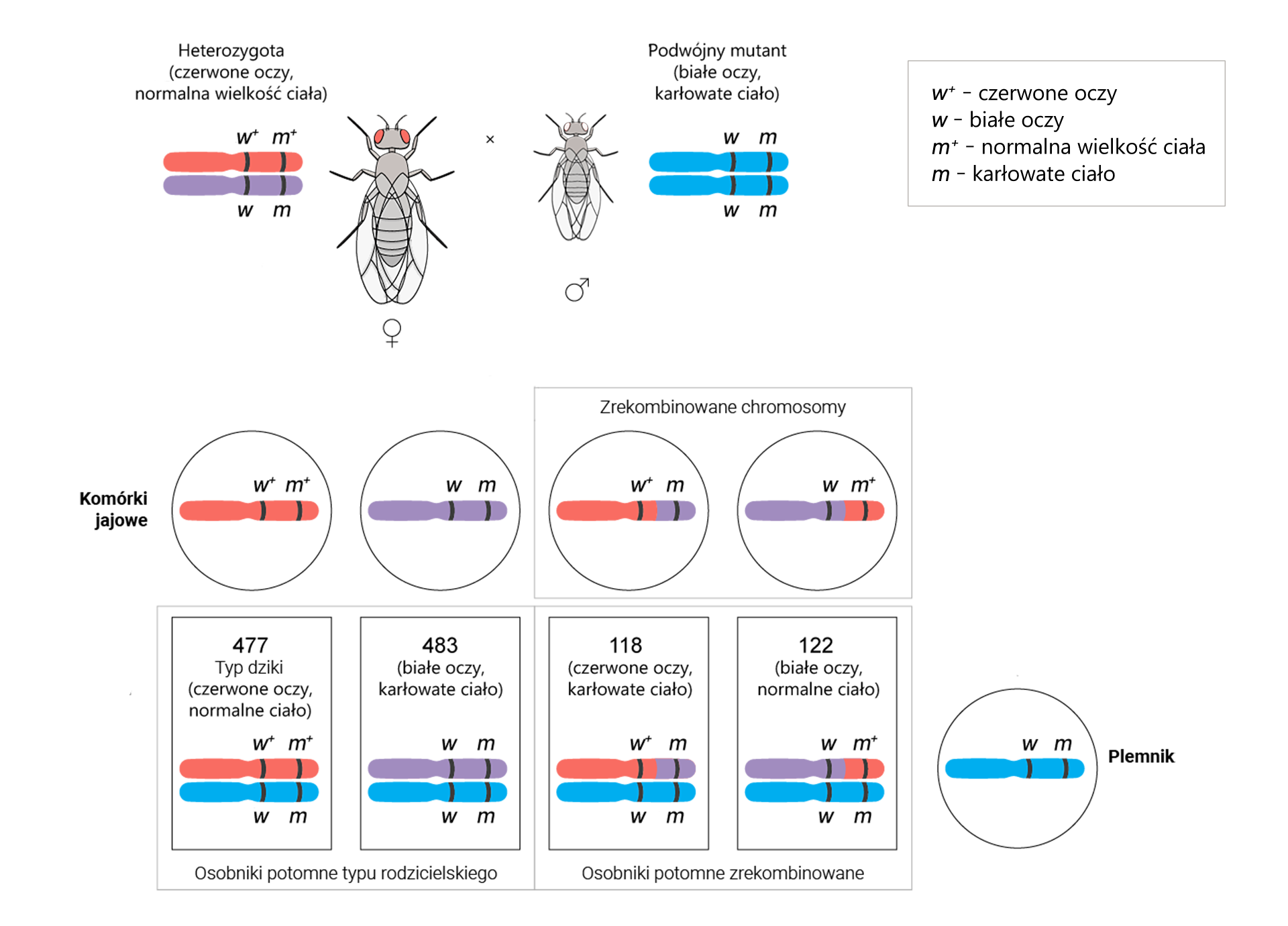
Geny , i u muszki owocowej (Drosophila melanogaster) są położone na tym samym chromosomie. Aby określić odległości pomiędzy nimi, wykonano krzyżówkę testową, czyli skrzyżowano podwójną heterozygotę / z podwójną homozygotą recesywną /.
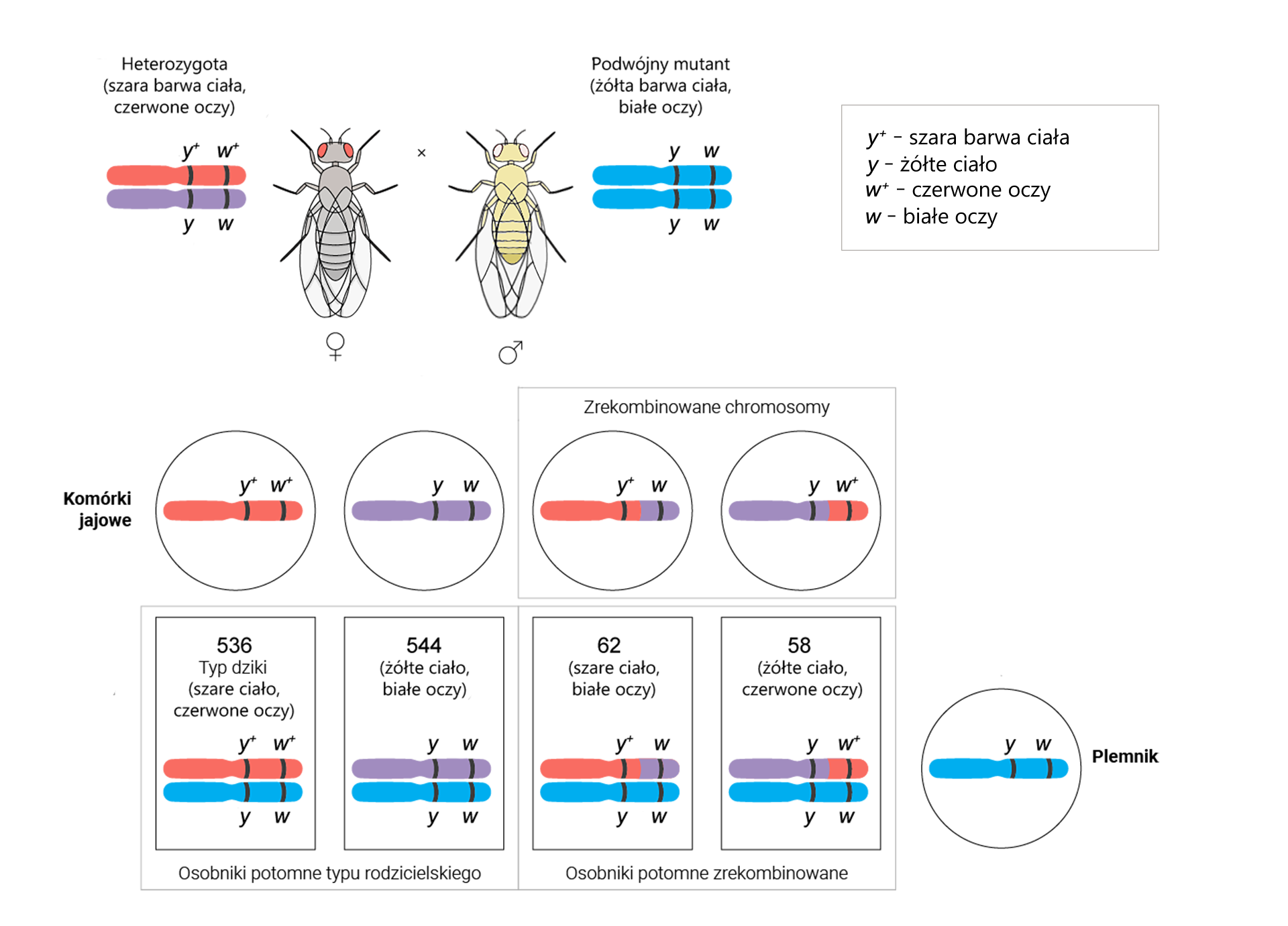
Otrzymano 1080 osobników o genach w układach rodzicielskich i 120 rekombinantów. Wartości te podstawiono do wzoru:
Odległość między genami = częstość rekombinacji = [liczba rekombinantów]/[całkowita liczba potomstwa] × 100% = [wynik]% = [wynik] cM
Odległość między genami i = [(62 + 58)/(536 + 544 + 62 + 58)] × 100% = (120/1200) × 100% = 10% = 10 cM
Z obliczeń wynika, że odległość między genami i wynosi 10 cM.
Następnie przeprowadzono analogiczne obliczenia w celu sprawdzenia odległości pomiędzy genami i .
W celu obliczenia odległości między genami i przeprowadzono następującą krzyżówkę testową. Na jej podstawie oblicz odległość między genami i .
Przeprowadzono również analogiczne obliczenia w celu sprawdzenia odległości pomiędzy genami i .
W celu obliczenia odległości między genami i przeprowadzono następującą krzyżówkę testową. Na jej podstawie oblicz odległość między genami i .
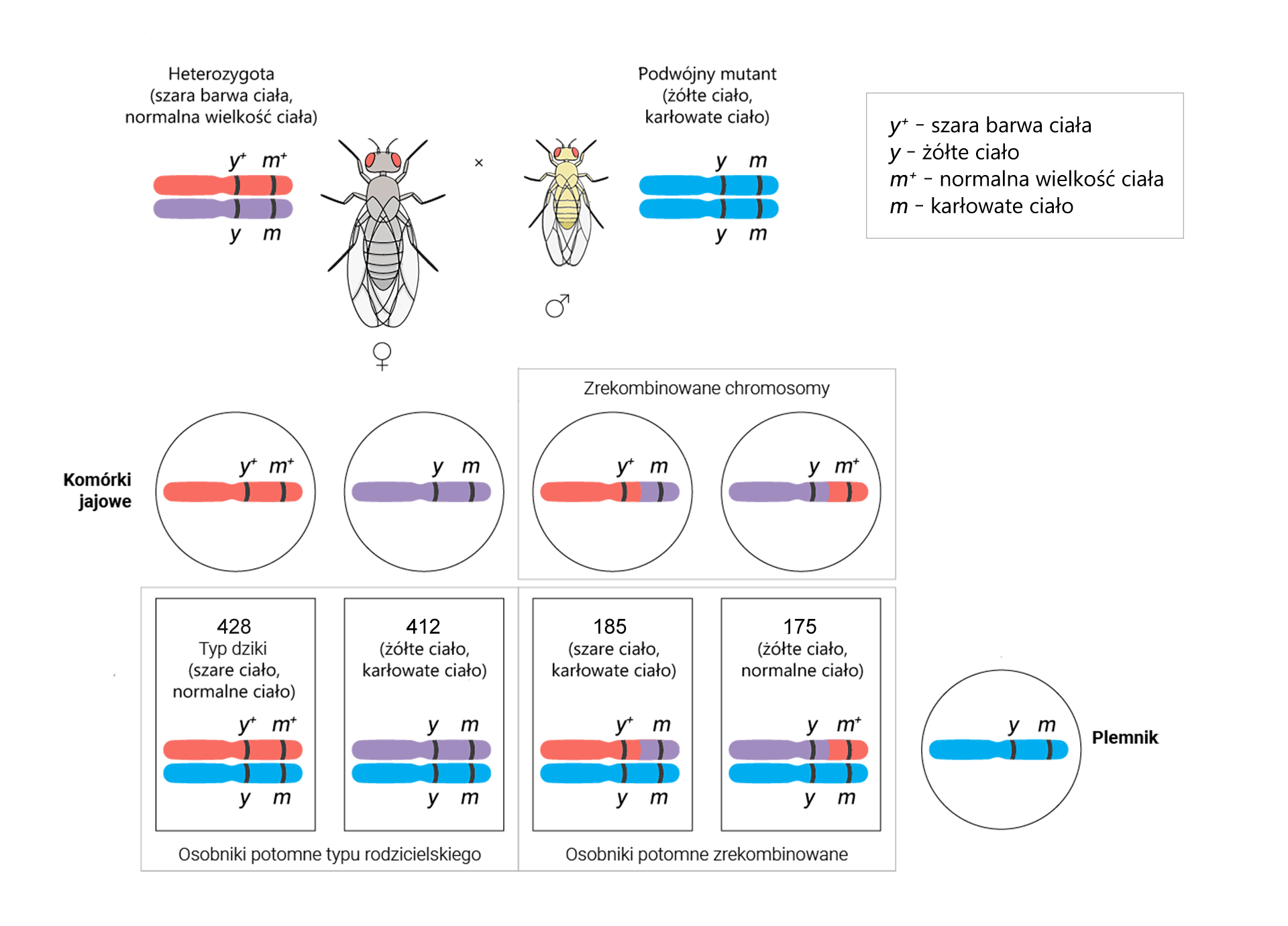
Znając częstości rekombinacji między poszczególnymi genami można ustalić ich kolejność na chromosomie.
Jeżeli gen znajduje się w odległości 10 cM od genu , gen w odległości 20 cM od genu , a gen 30 cM od genu , oznacza to, że na chromosomie gen znajduje się przed genem , a ten z kolei przed genem . Największa częstość rekombinacji oznacza, że geny i są najbardziej oddalone od siebie.
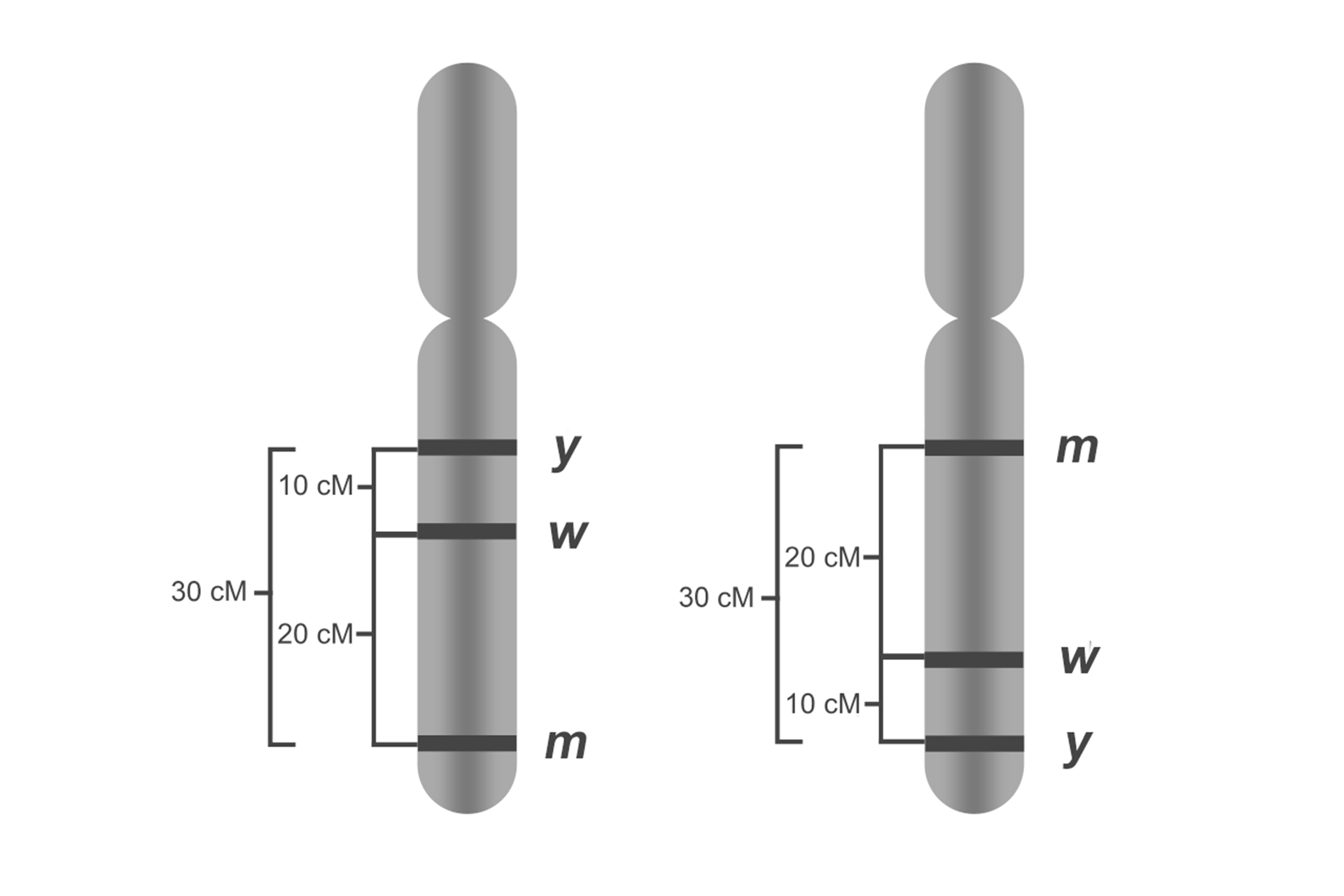
Odległość między genami mierzona za pomocą częstości rekombinantów pokazuje, jak geny są uszeregowane, nie pokazuje natomiast ich fizycznej odległości od siebie. Wynika to z faktu, że w pewnych punktach chromosomów znajdują się tzw. gorące miejsca rekombinacji, czyli takie miejsca, w których crossing‑over zachodzi częściej, niż wynikałoby to z teoretycznych obliczeń.
Aby poćwiczyć obliczanie odległości genami między genami na podstawie częstości występowania rekombinacji przeprowadź symulację:
Za pomocą symulacji interaktywnej oblicz częstości występowania rekombinacji pomiędzy poszczególnymi genami. Następnie określ odległości pomiędzy poszczególnymi genami, aby poznać ułożenie genów na chromosomie.
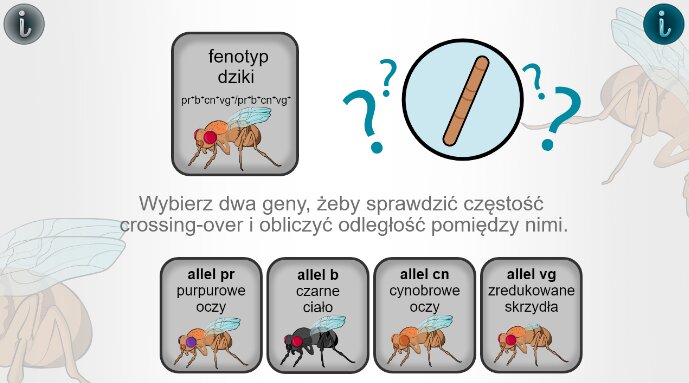
Zasób interaktywny dostępny pod adresem https://zpe.gov.pl/a/DD84M2KOH
Zapoznaj się z opisem symulacji:
Symulacja umożliwia obliczenie odległości miedzy czterema genami muszki owocowej na podstawie częstości występowania między nimi crossing‑over oraz ustalenie położenia tych genów na chromosomie. W początkowym menu można wybrać dwa z czterech genów: pr, b, cn i vg. Po wybraniu danej pary wyświetla się krzyżówka testowa podwójnej heterozygoty z podwójną homozygotą recesywną oraz w tabeli liczba osobników w potomstwie o układach rodzicielskich i o układach zrekombinowanych. Poniżej znajduje się wzór, do którego należy wpisać odczytane z tabeli wartości: częstość rekombinacji równa się liczba rekombinantów podzielić na całkowitą liczbę potomstwa razy sto procent, równa się wynik w procentach, równa się odległość między genami, równa się wynik w centymorganach. Należy po kolei wybrać wszystkie sześć par genów i prawidłowo wpisać wartości na podstawie krzyżówek, aby otrzymać wyniki: odległość pomiędzy genami b i pr wynosi 6 centymorganów, pomiędzy genami pr i cn wynosi 3 centymorgany, pomiędzy genami pr i vg wynosi 12,5 centymorgana, pomiędzy genami b i cn wynosi 9 centymorganów, pomiędzy genami b i vg wynosi 18,5 centymorganów, a odległość pomiędzy genami cn i vg wynosi 9,5 centymorgana. Po uzyskaniu wszystkich wyników wyświetla się ilustracja chromosomu na której trzeba umieścić fragmenty chromosomu o uzyskanych odległościach w ten sposób, aby wszystkie geny się pokrywały. W ten sposób powstaje chromosom z genami ułożonymi w kolejności: b, pr, cn i vg.
Mapowanie genów człowieka
Niektóre organizmy można łatwo hodować i rozmnażać w warunkach laboratoryjnych, np. muszkę owocową. W przypadku organizmów, które mają długi cykl życiowy albo których nie można hodować z przyczyn etycznych, np. człowieka, analizuje się ich rodowodyrodowody genetyczne. Są to graficzne przedstawienia informacji uzyskanych z wywiadu rodzinnego. Pozwalają na ustalenie występowania w rodzinie określonych cech, np. chorób genetycznych, i sposobów ich dziedziczenia. Analiza rodowodów zwykle poprzedza szczegółowe badania z wykorzystaniem technik biologii molekularnej.
Mapowanie genów pozwala naukowcom określić ich położenie na chromosomie, a zatem ocenić prawdopodobieństwo, że będą one dziedziczone wspólnie. Jest to bardzo użyteczne w hodowli zwierząt i roślin, pozwala bowiem na uzyskanie osobników o określonych pożądanych cechach i selekcję najlepszych z nich.
Podsumowanie
Badania Thomasa Morgana na muszce owocowej potwierdziły chromosomową teorię dziedziczenia:
- Chromosomy są liniowymi strukturami zawierającymi geny, które są ułożone liniowo i nie zachodzą na siebie.
- Gen zajmuje określone miejsce na chromosomie – locus; allele tego samego genu leżą w tym samym locus na chromosomach homologicznych.
- Geny znajdujące się na jednym chromosomie są sprzężone i mają tendencję do wspólnego dziedziczenia.
- Geny niesprzężone leżą na różnych (niehomologicznych) chromosomach i dziedziczą się niezależnie (II prawo Mendla).Geny sprzężone może rozdzielać m.in. proces crossing‑over, który prowadzi do powstawania rekombinantów (nowych układów alleli).
Krzyżówka testowa pozwala wykryć crossing‑over dzięki ujawnieniu fenotypów rodzicielskich (liczniejsze) i rekombinantów (mniej liczne).
Częstość crossing‑over oblicza się na podstawie odsetka rekombinantów i wyraża się w procentach.
Mapowanie genów pozwala ustalić kolejność genów i ich względne odległości na chromosomie. Jednostką odległości odległości jest centymorgan (cM).
Ćwiczenia utrwalające
Sprzężenie genów oznacza się zazwyczaj z wykorzystaniem… Możliwe odpowiedzi: 1. kreski ułamkowej, 2. dwukropka, 3. średnika, 4. dywizu
Uporządkuj w odpowiedniej kolejności geny , i na podstawie poniższej tabeli, zaczynając od genu B.
Geny | Częstość crossing‑over [%] |
2 | |
18 | |
20 |
Wróć do polecenia na stronie „Na dobry początek” i dopisz brakujące definicje. Pamiętaj, żeby nie kopiować słownika, ale wyjaśnić każde słowo kluczowe w miarę możliwości swoimi słowami.