Przeczytaj
Większość genów charakteryzuje się stałą, pełną ekspresywnościąekspresywnością – to oznacza, że wszystkie osobniki posiadające dany układ allelialleli wykazują ten sam fenotypfenotyp. Zdarza się jednak, że współdziałające ze sobą geny, oddziałują na siebie w taki sposób, że następuje zmiana ekspresywności. Przykładem zmiany ekspresywności genu jest epistaza.
Epistaza
Epistaza to interakcja dwóch genów położonych na różnych chromosomachchromosomach, w której gen epistatycznygen epistatyczny maskuje efekt ekspresji genu hipostatycznegogenu hipostatycznego.
W niektórych przypadkach działanie genu epistatycznego jest bardzo silne, dochodzi wówczas do nieprzejawiania się cechy uwarunkowanej przez gen hipostatyczny. Gen maskowany staje się nieczynny, dlatego że gen maskujący całkowicie tłumi jego obecność. Zjawisko to nazywa się hipostazą.
Przykład epistazy
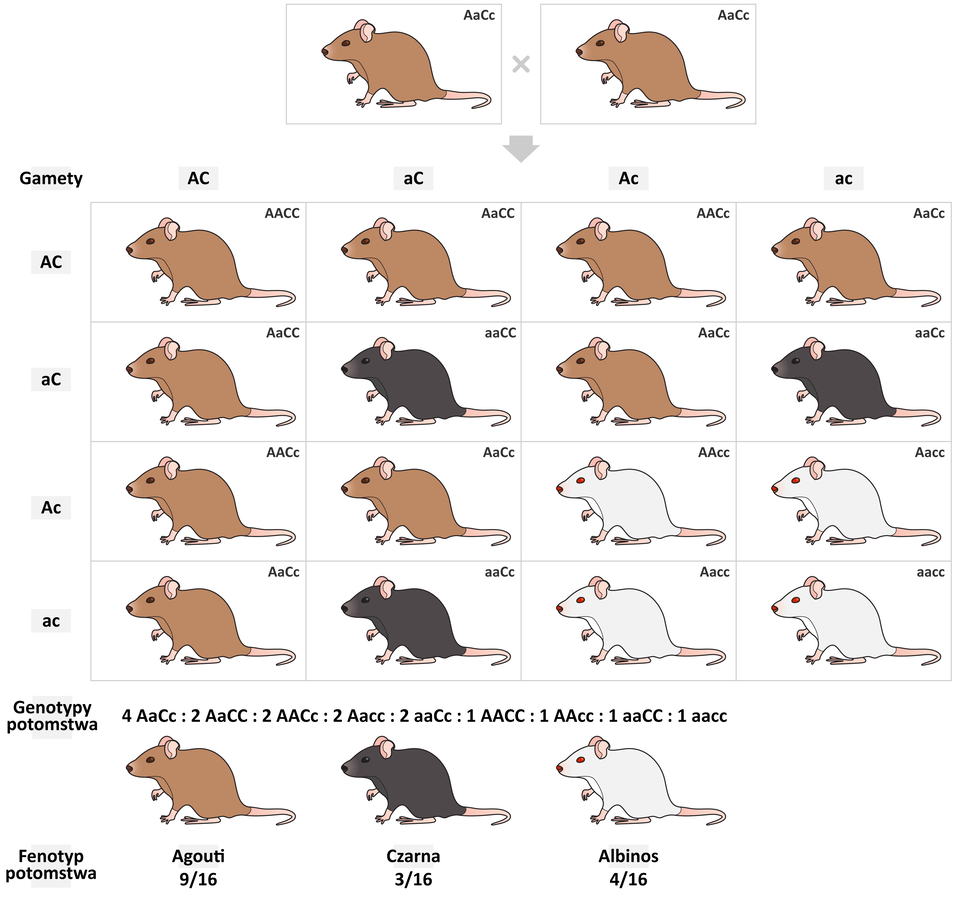
Przykładem epistazy pojedynczej recesywnej jest dziedziczenie barwy sierści u gryzoni, które wynika ze współdziałania dwóch różnych genów:
Gen A (zapisane dużą literą), wpływający na rozmieszczenie barwnika w strukturze włosa
duże A – allel warunkujący umaszczenie agouti
małe a – allel warunkujący umaszczenie jednolite
Gen C (zapisane dużą literą), wpływający na syntezę barwnika w strukturze włosa
duże C – allel odpowiadający za syntezę barwnika
małe c – allel odpowiadający za brak syntezy barwnika
Barwa sierści gryzoni zależy od występowania barwnika i jego rozmieszczenia w strukturze włosa. Obecność w genotypie przynajmniej jednego allelu C jest warunkiem niezbędnym do syntezy barwnika. Zatem osobniki o genotypie:
Barwa sierści gryzoni zależy od występowania barwnika i jego rozmieszczenia w strukturze włosa. Obecność w genotypie przynajmniej jednego allelu C (zapisanego dużą literą) jest warunkiem niezbędnym do syntezy barwnika. Zatem osobniki o genotypie:

Brak obecności allelu C w genotypie osobnika sprawia, że barwnik nie jest syntetyzowany. W takim przypadku układ alleli warunkujących rozmieszczenie barwnika w strukturze włosa nie ma znaczenia. Niezależnie od układu alleli genu A, takie osobniki mają zawsze białą sierść. Zatem osobniki o genotypie:
Brak obecności allelu C (zapisane dużą literą) w genotypie osobnika sprawia, że barwnik nie jest syntetyzowany. W takim przypadku układ alleli warunkujących rozmieszczenie barwnika w strukturze włosa nie ma znaczenia. Niezależnie od układu alleli genu A (zapisane dużą literą), takie osobniki mają zawsze białą sierść. Zatem osobniki o genotypie:
aacc, Aacc, AAcc są albinotyczne
dwa małe a dwa małe c, duże A małe a dwa małe c, dwa duże A dwa małe c są albinotyczne
Allel c jest genem epistatycznym, czyli nadrzędnym w stosunku do alleli A i a. Allele A i a są hipostatyczne, czyli podrzędne w stosunku do allelu c. Zatem za fenotypowe ujawnienie się genu hipostatycznego, odpowiada gen epistatyczny.
Allel c (małą literą) jest genem epistatycznym, czyli nadrzędnym w stosunku do alleli duże A i małe a. Allele duże A i małe a są hipostatyczne, czyli podrzędne w stosunku do allelu małe c. Zatem za fenotypowe ujawnienie się genu hipostatycznego, odpowiada gen epistatyczny.
Rozkład cech w przypadku epistazy pojedynczej recesywnej przedstawiono na przykładzie dziedziczenia barwy sierści u gryzoni. Organizmy rodzicielskie to podwójne heterozygoty (AaCc) o barwie agouti. Osobniki posiadające przynajmniej po jednym allelu dominującym obu genów będą miały barwę agouti. U osobników, które będą homozygotą recesywną genu A i będą posiadały chociaż jeden allel dominujący genu C barwa sierści będzie czarna. Organizmy homozygotyczne recesywne genu C będą barwy białej, niezależnie od układu alleli genu A.
Rozkład cech w przypadku epistazy pojedynczej recesywnej przedstawiono na przykładzie dziedziczenia barwy sierści u gryzoni. Organizmy rodzicielskie to podwójne heterozygoty (duże A małe a duże C małe c) o barwie agouti. Osobniki posiadające przynajmniej po jednym allelu dominującym obu genów będą miały barwę agouti. U osobników, które będą homozygotą recesywną genu duże A i będą posiadały chociaż jeden allel dominujący genu duże C barwa sierści będzie czarna. Organizmy homozygotyczne recesywne genu duże C będą barwy białej, niezależnie od układu alleli genu duże A.
Przykładem epistazy pojedynczej recesywnej jest także dziedziczenie barwy sierści u psów Labrador retriever. Umaszczenie może mieć barwę czarną, brązową lub biszkoptową.
U ludzi przykładem epistazy jest dziedziczenie barwy skóry. Cecha ta determinowana jest przez wiele genów. Jednak to od jednego genu, odpowiedzialnego za syntezę enzymu – tyrozynazytyrozynazy, zależy działanie pozostałych genów warunkujących barwę skóry. Funkcjonalna forma tyrozynazy jest niezbędna w biosyntezie melaninymelaniny. Skutkiem mutacji genu kodującego enzym, jest brak tyrozynazy, w wyniku czego barwnik nie jest wytwarzany. Skóra, włosy i tęczówka oka są jasne, a więc taki osobnik jest albinosem. Zatem gen kodujący enzym jest episatyczny wobec pozostałych genów kodujących barwnik.
Rodzaje epistazy
Zgodnie z drugim prawem Mendla, allele dwóch różnych par genów rozdzielane są do gamet na zasadzie losowej segregacji. Efektem skrzyżowania dwóch osobników heterozygotycznych względem obu genów (AaBb) jest rozkład cech 9:3:3:1, przy dziewięciu różnych genotypach o rozkładzie 1:1:2:2:4:2:2:1:1. W przypadku epistazy, stosunek genotypowy pozostaje niezmienny, lecz stosunek fenotypowy ulega zmianie.
Zgodnie z drugim prawem Mendla, allele dwóch różnych par genów rozdzielane są do gamet na zasadzie losowej segregacji. Efektem skrzyżowania dwóch osobników heterozygotycznych względem obu genów (duże A małe a duże B małe b) jest rozkład cech 9:3:3:1, przy dziewięciu różnych genotypach o rozkładzie 1:1:2:2:4:2:2:1:1. W przypadku epistazy, stosunek genotypowy pozostaje niezmienny, lecz stosunek fenotypowy ulega zmianie.
Więcej informacji o drugim prawie Mendla oraz segregacji znajdziesz w materiale: Prawo niezależnego dziedziczenia cechPrawo niezależnego dziedziczenia cech.
Relacje epistatyczne, mogą mieć różne formy, które charakteryzują się odmiennymi rozkładami fenotypów:
epistaza pojedyncza dominująca, kiedy gen epistatyczny jest dominujący; obserwowany stosunek fenotypów w dziedziczeniu dwugenowym to 12:3:1 - np. barwa cukinii
epistaza pojedyncza recesywna, kiedy gen epistatyczny jest recesywny; obserwowany stosunek fenotypów w dziedziczeniu dwugenowym to 9:3:4 - np. barwa sierści labradorów
epistaza podwójna dominująca, kiedy dwa geny kontrolujące tę samą reakcję są dominujące i tylko podwójna homozygota recesywna ujawni fenotyp recesywny; obserwowany stosunek fenotypów w dziedziczeniu dwugenowym to 15:1 - np. dziedziczenie kształtu ziarna pszenicy
epistaza podwójna recesywna, kiedy proces biochemiczny jest katalizowany przez produkty dwóch różnych genów, czyli obie reakcje muszą zajść, aby cecha się ujawniła; obserwowany stosunek fenotypów w dziedziczeniu dwugenowym to 9:7 - np. barwa kwiatów grochu pachnącego
- Nazwa kategorii: Rodzaje epistazy
- Nazwa kategorii: epistaza pojedyncza
- Nazwa kategorii: dominująca
- Nazwa kategorii: recesywna Koniec elementów należących do kategorii epistaza pojedyncza
- Nazwa kategorii: epistaza podwójna
- Nazwa kategorii: recesywna
- Nazwa kategorii: dominująca Koniec elementów należących do kategorii epistaza podwójna
- Elementy należące do kategorii Rodzaje epistazy
- Elementy należące do kategorii epistaza pojedyncza
- Elementy należące do kategorii epistaza podwójna
Słownik
rodzaj umaszczenia ssaków, powstaje na skutek naprzemiennego rozmieszczenia dwóch barwników
forma genu; jedna z wersji genu różniąca się od pozostałych sekwencją nukleotydów; zajmuje określone miejsce na chromosomie
(gr. chrṓma – barwa, sṓma – ciało) forma organizacji materiału genetycznego, pojawiająca się w czasie mitozy i mejozy; najwyższy stopień upakowania chromatyny, umożliwiający równy podział informacji genetycznej komórkom potomnym
stopień w jakim cecha uwarunkowana obecnością danego układu alleli ujawnia się w fenotypie osobnika
gen wpływający na wyrażenie się efektu fenotypowego innego genu (hipostatycznego)
(gr. phaínomai – pojawiam się, týpos – odbicie) zespół anatomicznych, fizjologicznych i biochemicznych cech organizmu uwarunkowanych współdziałaniem genotypu i warunków środowiska
gen, którego efekt fenotypowy zależy od współdziałania genu epistatycznego
barwnik nadający kolor skórze, włosom i tęczówce oka; chroni przed szkodliwym wpływem promieniowania UV
enzym katalizujący reakcję przekształcania tyrozyny do
3,4‑dihydroksyfenyloalaniny, związku wykorzystywanego do syntezy melanin