Przeczytaj

Im większa różnorodność genetyczna populacji, tym mniejsze ryzyko jej wyginięcia na skutek wystąpienia jakiegoś niekorzystnego czynnika. Zróżnicowanie puli genowej populacji daje bowiem szansę, że część osobników przeżyje dzięki obecności u nich zestawu genówgenów zwiększających przystosowanie organizmu do zmieniającego się środowiska.
Mała różnorodność genetyczna populacji może być skutkiem kojarzenia się osobników blisko ze sobą spokrewnionych – tzw. kojarzenia krewniaczego. Zwiększa ono prawdopodobieństwo dziedziczenia od obojga rodziców alleli recesywnych i tym samym powstawania homozygot recesywnych, co powoduje ujawnianie się w fenotypie niekorzystnych dla organizmu cech uwarunkowanych allelami recesywnymi.
Crossing‑over

Crossing‑overCrossing‑over jest jednym z mechanizmów rekombinacjirekombinacji. Proces ten polega na wymianie fragmentów chromatyd pomiędzy dwoma chromosomami homologicznymi podczas profazy pierwszego podziału mejotycznego. Może zajść w różnych miejscach chromosomu, doprowadzając do wymiany mniejszego (gdy wymiana zajdzie bliżej końca ramienia chromosomu) lub większego fragmentu chromosomu (gdy nastąpi bliżej centromerucentromeru).
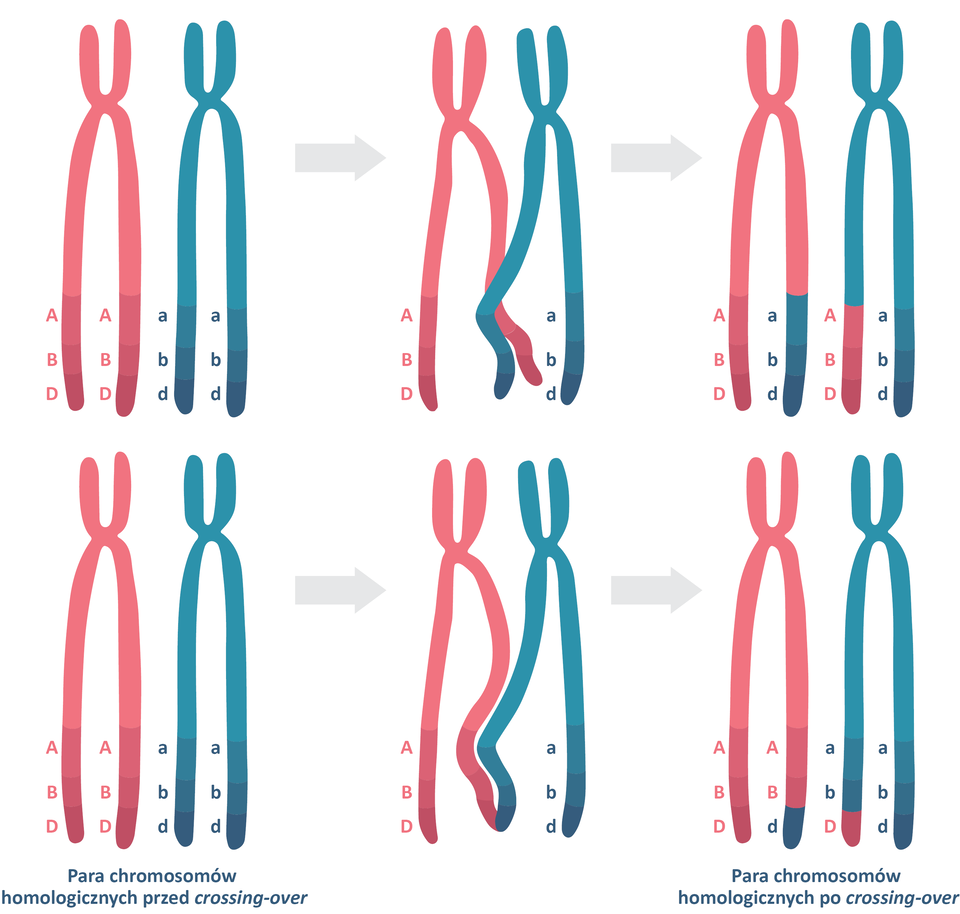
Wyobraźmy sobie chromosom i zaznaczmy na nim dwa hipotetyczne geny: jeden położony blisko końca ramienia, a drugi blisko centromeru. Pomiędzy ich loci jest duża odległość, dlatego istnieje wysokie prawdopodobieństwo, że podczas mejozy komórki macierzystej ich allele zostaną rozdzielone na skutek crossing‑over i trafią do gamet w innej konfiguracji niż w układach rodzicielskich. W związku z tym powstaną cztery rodzaje gamet, w których wystąpią zarówno rodzicielskie układy genów, jak i nowe kombinacje alleli.
Gdy geny sprzężone leżą blisko siebie na jednym chromosomie, zwykle trafiają do gamet razem, ponieważ prawdopodobieństwo ich rozdzielenia podczas crossing‑over jest niewielkie. W związku z tym najczęściej powstają gamety zawierające rodzicielskie układy alleli genów, a znacznie rzadziej – gamety zawierające układy zrekombinowane na skutek crossing‑over.
Obliczanie częstości crossing‑over
Na podstawie liczby rekombinantów można określić częstość zachodzenia zjawiska crossing‐over pomiędzy dwoma genami na chromosomie. Obliczeń tych dokonuje się na podstawie wyników krzyżówki testowej.
Przeprowadza się je następująco:
Poniżej przedstawiono przykład obliczeń częstości crossing‑over pomiędzy genami b i vg u muszki owocowej. Allele warunkujące wystąpienie cech typu dzikiego zapisuje się ze znakiem „+”, natomiast allele odpowiadające za wystąpienie mutacji - bez plusa. Zapis bIndeks górny ++oznacza więc allel warunkujący szarą barwę ciała (fenotyp dziki), natomiast allel b (od angielskiego black, czyt. blak) – allel warunkujący wystąpienie mutacji objawiającej się czarną barwą ciała. Allel warunkujący skrzydła normalnej długości zapisuje się jako vgIndeks górny ++, a allel objawiający się zredukowanymi skrzydłami vg (od angielskiego vestigial, czyt. westidżial).
Obliczenie częstości crossing‑over wymaga wielu analiz statystycznych na dużej liczbie badanych osobników. Wartość tę podaje się w procentach. Na przykład crossing‑over wynoszące 10% oznacza, że u 10% potomstwa nastąpi rozdzielenie sprzężonych ze sobą alleli badanych genów.
Obliczanie odległości między genami oraz określenie kolejności ich położenia na chromosomie
Znając częstość zachodzenia crossing‑over na konkretnym odcinku, tzn. między dwoma konkretnymi genami, można obliczyć zarówno odległość między poszczególnymi genami na chromosomie, jak i określić kolejność ich położenia.
Amerykański biolog Thomas Morgan, który opracował chromosomową teorię dziedziczenia cech, został upamiętniony w nazwie jednostki odległości między genami: 1 centymorgan (cM), inaczej 1 jednostka mapowa (j.m.), odpowiada 1% częstości crossing‑over.
Graficzne przedstawienie umiejscowienia poszczególnych genów na chromosomie nazywa się mapą genetyczną, a obliczenia – mapowaniem chromosomu lub mapowaniem genów.
Przykład obliczania odległości między poszczególnymi genami na chromosomie oraz ich kolejności położenia:
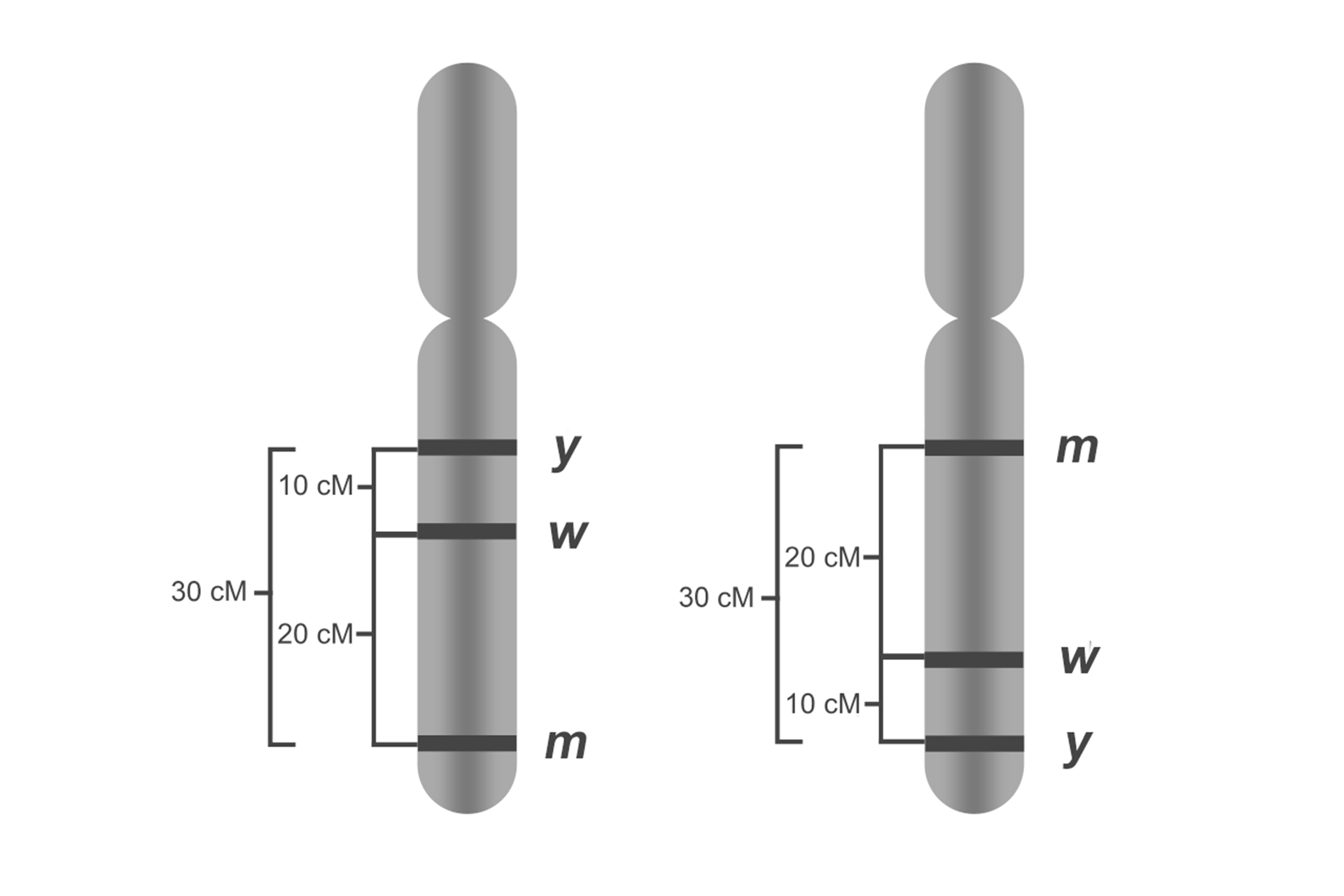
Geny y, w i m u muszki owocowej (Drosophila melanogaster) są położone na tym samym chromosomie. Aby określić odległości pomiędzy nimi, wykonano krzyżówkę testowąkrzyżówkę testową, czyli skrzyżowano podwójną heterozygotę yIndeks górny ++wIndeks górny ++/yw z podwójną homozygotą recesywną yw/yw.
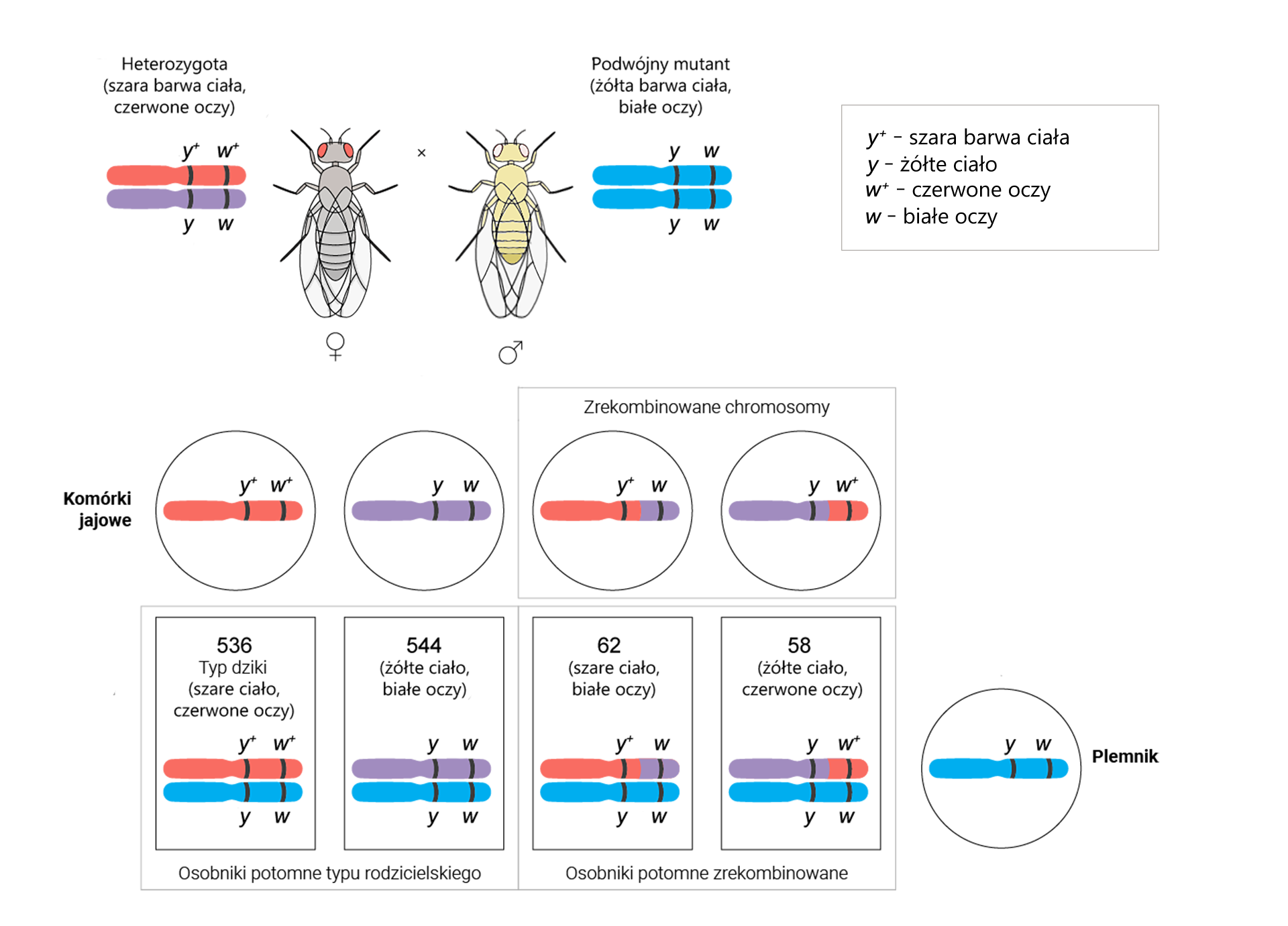
Otrzymano 1080 osobników o genach w układach rodzicielskich i 120 rekombinantów. Wartości te podstawiono do wzoru:
Odległość między genami = częstość rekombinacji = [liczba rekombinantów]/[całkowita liczba potomstwa] × 100% = [wynik]% = [wynik] cM
Odległość między genami y i w = [(62 + 58)/(536 + 544 + 62 + 58)] × 100% = (120/1200) × 100% = 10% = 10 cM
Z obliczeń wynika, że odległość między genami y i w wynosi 10 cM.
Następnie przeprowadzono analogiczne obliczenia w celu sprawdzenia odległości pomiędzy genami w i m.
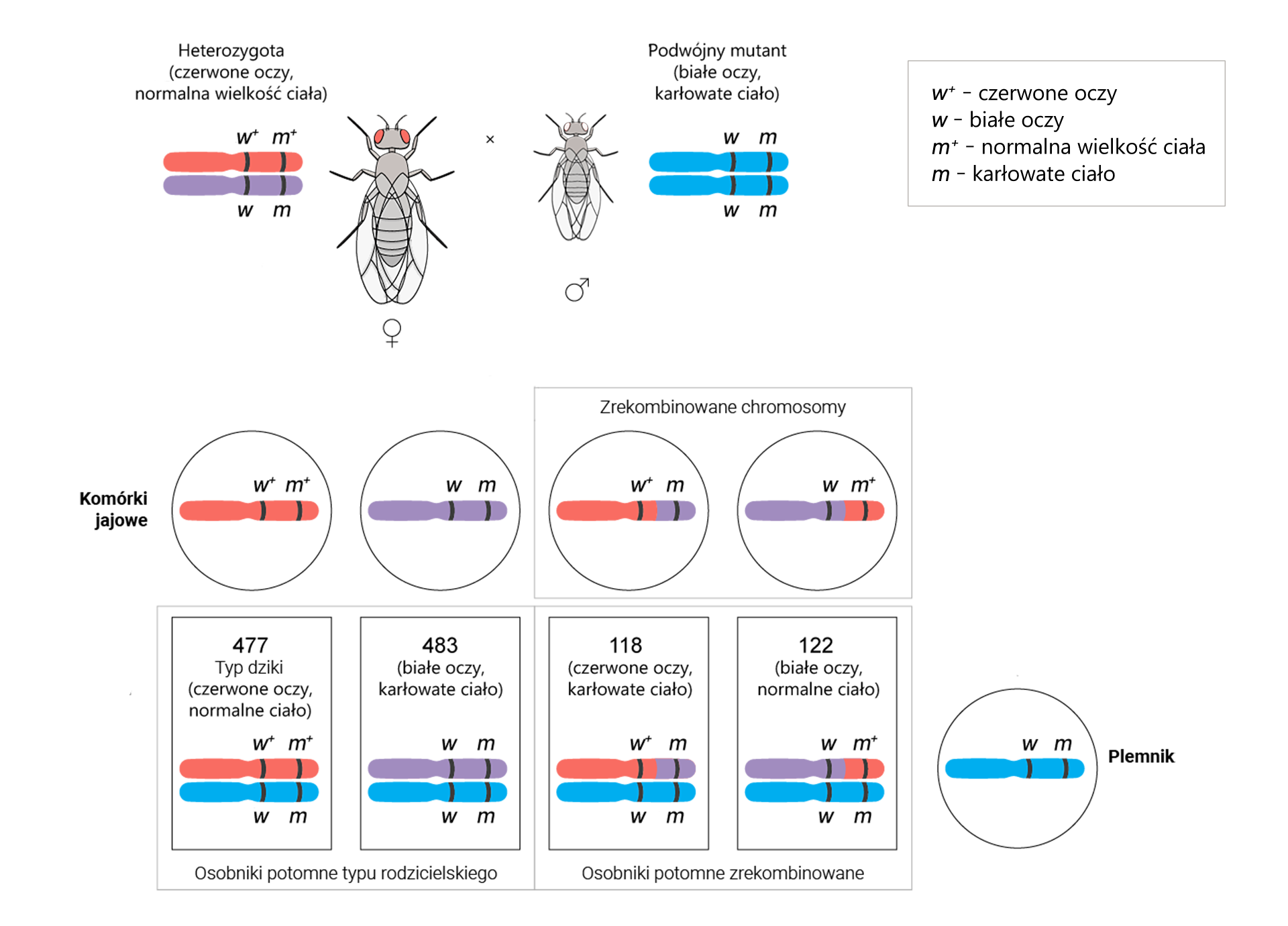
Otrzymano 960 osobników o genach w układach rodzicielskich i 240 rekombinantów. Wartości te podstawiono do wzoru.
Odległość między genami w i m = [(118 + 122)/(477 + 483 + 118 + 122)] × 100% = (240/1200) × 100% = 20% = 20 cM
Przeprowadzono również analogiczne obliczenia w celu sprawdzenia odległości pomiędzy genami y i m.
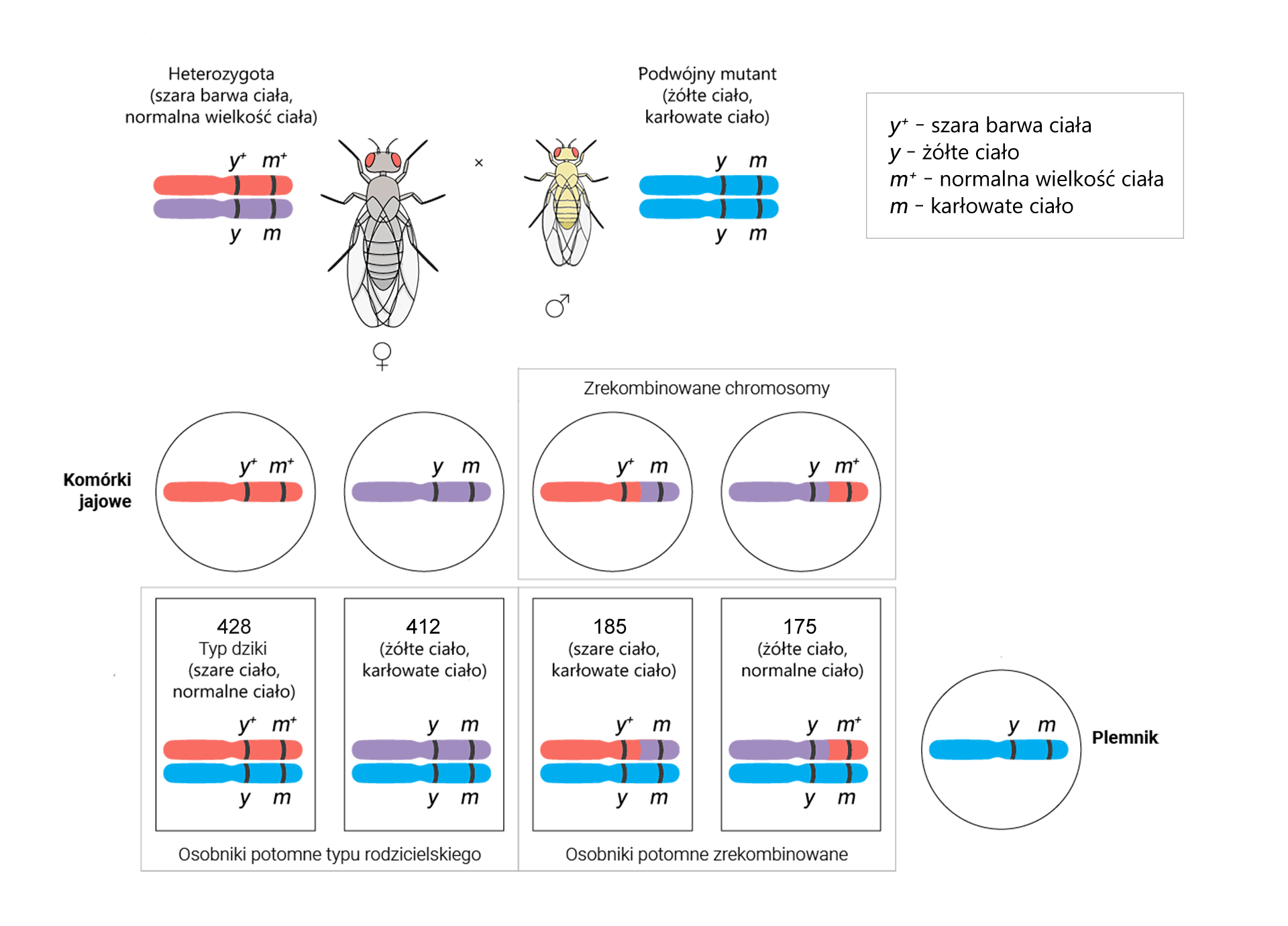
Otrzymano 840 osobników o genach w układach rodzicielskich i 360 rekombinantów. Wartości te podstawiono do wzoru.
Odległość między genami y i m = [(185 + 175)/(428 + 412 + 185 + 175)] × 100% = (360/1200) × 100% = 30% = 30 cM
Znając częstości rekombinacji między poszczególnymi genami można ustalić ich kolejność na chromosomie.
Jeżeli gen y znajduje się w odległości 10 cM od genu w, gen w w odległości 20 cM od genu m, a gen y 30 cM od genu m, oznacza to, że na chromosomie gen y znajduje się przed genem w, a ten z kolei przed genem m. Największa częstość rekombinacji oznacza, że geny y i m są najbardziej oddalone od siebie.
Odległość między genami mierzona za pomocą częstości rekombinantów pokazuje, jak geny są uszeregowane, nie pokazuje natomiast ich fizycznej odległości od siebie. Wynika to z faktu, że w pewnych punktach chromosomów znajdują się tzw. gorące miejsca rekombinacji, czyli takie miejsca, w których crossing‑over zachodzi częściej, niż wynikałoby to z teoretycznych obliczeń.
Podwójne crossing‑over
Dodatkowe utrudnienie dla obliczeń stanowi fakt, że crossing‑over może zajść dwukrotnie w obrębie jednej pary chromosomów. Zjawisko to określa się podwójnym crossing‑over. Na poniższym przykładzie w przypadku badania jedynie genów A i D podwójne crossing‑over jest niewidoczne, ponieważ geny te wracają do takiego ułożenia jak w układzie rodzicielskim.
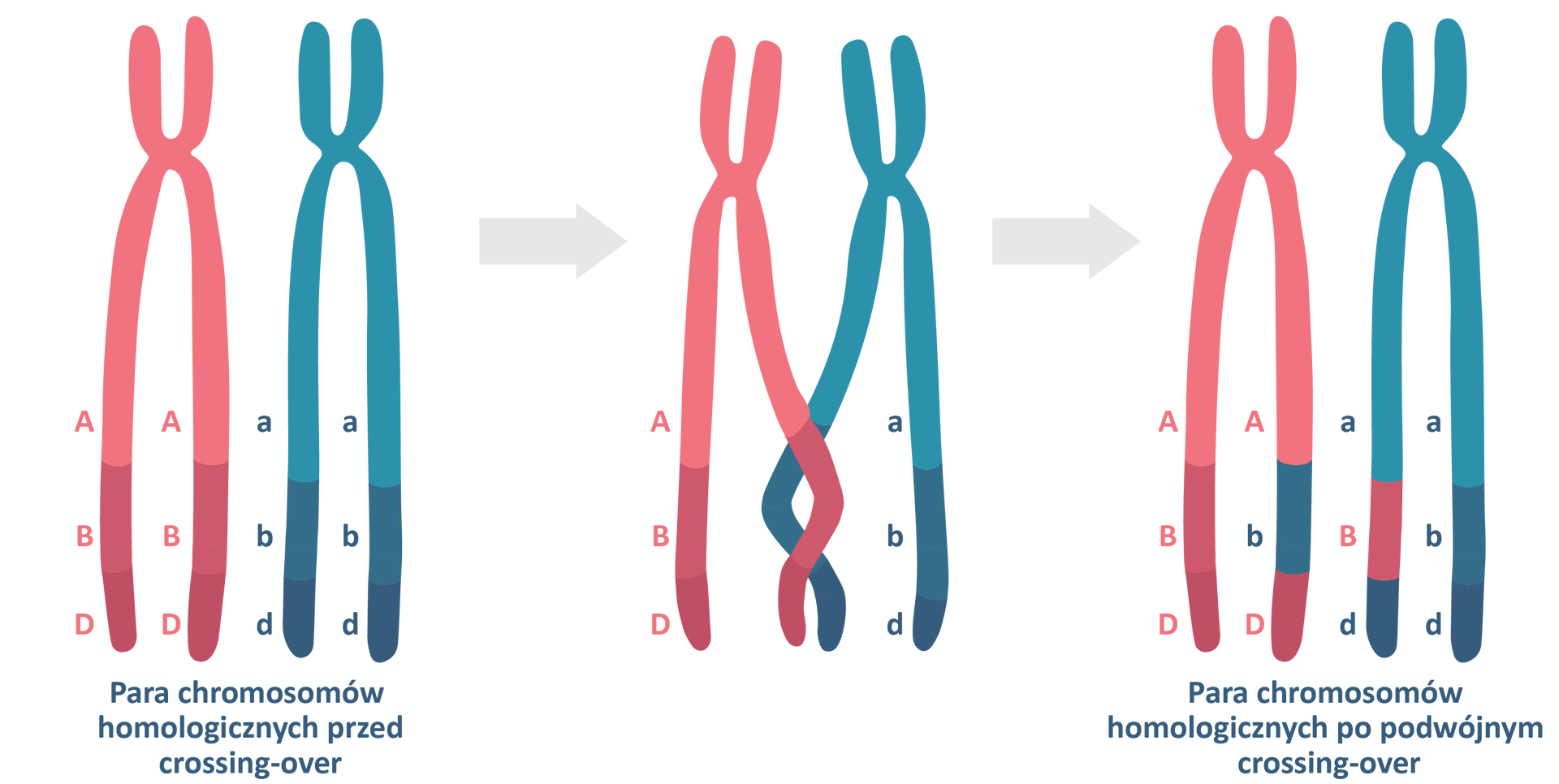
Z tego powodu, mapując położenie genów na chromosomach, bada się wiele osobników z wielu pokoleń. Im więcej obiektów w badaniu statystycznym, tym jest ono dokładniejsze, choć nigdy nie daje 100‑procentowo pewnych wyników.
Mapowanie genów człowieka
Niektóre organizmy można łatwo hodować i rozmnażać w warunkach laboratoryjnych, np. muszkę owocową. W przypadku organizmów, które mają długi cykl życiowy albo których nie można hodować z przyczyn etycznych, np. człowieka, analizuje się ich rodowodyrodowody genetyczne. Są to graficzne przedstawienia informacji uzyskanych z wywiadu rodzinnego. Pozwalają na ustalenie występowania w rodzinie określonych cech, np. chorób genetycznych, i sposobów ich dziedziczenia. Analiza rodowodów zwykle poprzedza szczegółowe badania z wykorzystaniem technik biologii molekularnej.
Mapowanie genów pozwala naukowcom określić ich położenie na chromosomie, a zatem ocenić prawdopodobieństwo, że będą one dziedziczone wspólnie. Jest to bardzo użyteczne w hodowli zwierząt i roślin, pozwala bowiem na uzyskanie osobników o określonych pożądanych cechach i selekcję najlepszych z nich.
Słownik
miejsce połączenia dwóch chromatyd siostrzanych; do niego przyłączają się włókna wrzeciona podziałowego, przez co stanowi część chromosomu odpowiedzialną za segregację chromosomów podczas podziału komórki (mitozy i mejozy)
proces zachodzący podczas profazy I podziału mejotycznego jądra komórkowego
fragment DNA (lub u niektórych wirusów RNA) kodujący określone białko lub RNA
krzyżówka genetyczna osobnika o fenotypie dominującym i nieznanym genotypie z homozygotą recesywną; pozwala ustalić, czy testowany osobnik jest homozygotą dominującą czy heterozygotą
proces prowadzący do powstawania nowych układów alleli genów; nie prowadzi do powstawania nowych genów; zachodzi na skutek niezależnej segregacji chromosomów podczas I podziału mejotycznego, crossing‑over oraz losowego łączenia się gamet podczas zapłodnienia
zestawienie genealogiczne obejmujące męskich i żeńskich przodków danej osoby, niekiedy w formie schematu graficznego, tzw. drzewa genealogicznego