Przeczytaj
W cyklach rozwojowych protistów roślinopodobnych, grzybów i roślin występuje przemiana pokoleń połączona z przemianą faz jądrowychprzemianą faz jądrowych.
Przemiana pokoleń
Przemiana pokoleń to zjawisko regularnego następowania po sobie pokolenia rozmnażającego się płciowo przy użyciu gamet – gametofitugametofitu i pokolenia rozmnażającego się bezpłciowo przy użyciu spor – sporofitusporofitu. Gametofit wytwarza gametangiagametangia, w których na drodze mitozy powstają haploidalne gamety: w plemniachplemniach – plemniki, a w lęgniachlęgniach lub rodniachrodniach – komórki jajowe. W procesie zapłodnienia (gamii) haploidalne gamety łączą się ze sobą, w wyniku czego powstaje diploidalna zygota, z której rozwija się sporofit. Dojrzały sporofit wytwarza sporangia (zarodniezarodnie), w których na drodze mejozy powstają haploidalne spory (zarodniki), z nich zaś wyrastają gametofity.
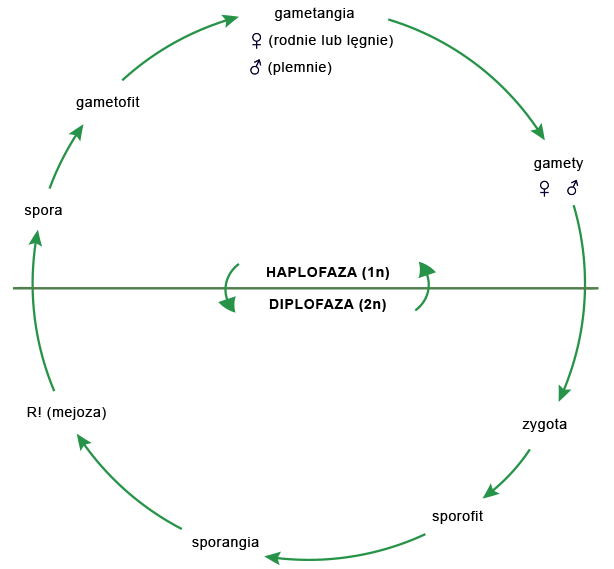
Typy przemiany pokoleń
Wyróżnia się dwa zasadnicze typy przemiany pokoleń: izomorficzną i heteromorficzną. W izomorficznej przemianie pokoleń gametofit i sporofit nie wykazują różnic morfologicznych. Występuje ona u niektórych protistów roślinopodobnych i roślin pierwotnie wodnych. Prawdopodobnie typ ten występował także u wymarłych pierwszych roślin lądowych. W heteromorficznej przemianie pokoleń gametofit i sporofit różnią się morfologicznie. Występuje ona u niektórych protistów roślinopodobnych oraz współcześnie żyjących roślin lądowych i wtórnie wodnych. Jedynie mszaki są grupą roślin, których pokoleniem dominującym jest gametofit. U pozostałych grup, czyli paprotników jednakozarodnikowych i różnozarodnikowych oraz roślin nagonasiennych i okrytonasiennych, dominuje sporofit.

Gametofit i gametangia u mszaków, paprotników, roślin nagonasiennych i roślin okrytonasiennych
Gametofit i gametangia u mszaków
Gametofit i gametangia u paprotników jednakozarodnikowych
Gametofit i gametangia u paprotników różnozarodnikowych
Gametofit i gametangia u roślin nagonasiennych
Gametofit i gametangia u roślin okrytonasiennych
Sporofit i sporangia u mszaków, paprotników, roślin nagonasiennych i roślin okrytonasiennych
Sporofit i sporangium u mszaków
Sporofit i sporangia u paprotników jednakozarodnikowych
Sporofit i sporangia u paprotników różnozarodnikowych

Sporofit i sporangia u roślin nagonasiennych

Sporofit i sporangia u roślin okrytonasiennych

Tendencje ewolucyjne w przemianie pokoleń roślin
Rozwój sporofitu i redukcja gametofitu
W ewolucji roślin obserwuje się wyraźną tendencję do zwiększania udziału sporofitu w cyklu życiowym rośliny. Początkowo sporofit był tworem o stosunkowo prostej budowie, całkowicie zależnym od gametofitu. Współcześnie tę linię rozwojową prezentują mszaki – zależność niesamodzielnego, heterotroficznegoheterotroficznego, krótkotrwałego sporofitu od samodzielnego, autotroficznegoautotroficznego i wieloletniego gametofitu jest w tej grupie roślin lądowych bardzo wyraźna. Natomiast u wymarłych i współcześnie żyjących paprotników, roślin nagonasiennych i roślin okrytonasiennych zaznacza się tendencja do rozwoju i specjalizacji sporofitu, który stopniowo uniezależniał się od gametofitu, stając się okazałą, samodzielną, autotroficzną i wieloletnią rośliną o skomplikowanej budowie. Zdolność sporofitu do wykształcenia wyspecjalizowanych tkanek roślinnych i właściwych organów wegetatywnychorganów wegetatywnych umożliwiała lepsze przystosowanie się do warunków środowiska. Równolegle z postępującą dominacją sporofitu następowała stopniowa redukcja gametofitu, który stał się niesamodzielnym, kilkukomórkowym tworem, żyjącym w obrębie tkanek sporofitu i całkowicie od niego zależnym.
Potencjał rozwojowy sporofitu wynika z diploidalności komórek. Podwojony zestaw chromosomów powoduje, że każdy gen reprezentowany jest przez dwa alleleallele. Jeśli jeden z dwóch alleli danego genu zawiera niekorzystną mutację, to drugi prawidłowy allel zwiększa szansę na przeżycie organizmu. W przypadku haploidalnego gametofitu mutacja genu warunkującego cechę istotną dla przeżycia organizmu często oznacza jego śmierć.
Przejście z jednakozarodniowości do różnozarodnikowości
U pierwszych roślin zarodniki (spory), tworzone w dużych ilościach w zarodniach (sporangiach), były jednakowe pod względem morfologicznym i fizjologicznym. Wyrastały z nich gametofity jednopiennegametofity jednopienne, które wytwarzały zarówno rodnie, jak i plemnie. Taki rozwój zarodników wciąż występuje u części mszaków i paprotników jednakozarodnikowych. U niektórych z tych roślin pojawiło się zróżnicowanie fizjologiczne zarodników: z jednakowych morfologicznie zarodników wyrastają gametofity dwupiennegametofity dwupienne – żeńskie (wytwarzające rodnie) i męskie (wytwarzające plemnie). U paprotników różnozarodnikowych, roślin nagonasiennych i okrytonasiennych nastąpiło zróżnicowanie zarodników (spor) i zarodni (sporangiów): duże zarodniki (makrosporymakrospory) powstają w dużych zarodniach (makrosporangiachmakrosporangiach) i małe zarodniki (mikrosporymikrospory) powstają w małych zarodniach (mikrosporangiachmikrosporangiach). Zróżnicowaniu morfologicznemu towarzyszy także zróżnicowanie fizjologiczne: z makrospory wyrasta gametofit żeńskigametofit żeński, a z mikrospory wyrasta gametofit męskigametofit męski. Dalsze przekształcenia obejmowały przede wszystkim makrospory: stopniowo zmniejszała się liczba funkcjonalnych makrospor – do jednej u roślin nagonasiennych i okrytonasiennych, u których nie opuszcza ona już makrosporangium. Makrosporangium z rozwijającą się w jego wnętrzu makrosporą zostaje otoczone dodatkowymi osłonkami, co prowadzi do powstania zalążka.
Uniezależnienie procesu zapłodnienia od wody
Proces zapłodnienia u roślin odbywał się początkowo z udziałem wody, w której plemniki opatrzone wiciami przepływały z plemni do rodni. Współcześnie tę tendencję ewolucyjną prezentują mszaki i paprotniki jednako- oraz różnozarodnikowe, które z uwagi na taki proces zapłodnienia są nierozerwalnie związane z wilgotnymi siedliskami środowiska lądowego. Możliwość opanowania innych siedlisk była związana z udoskonaleniem procesu zapłodnienia, który stał się niezależny od obecności wody. Przenośnikiem gamet męskich stały się mikrospory, które transportowane przez wiatr, zwierzęta lub wodę docierają bezpośrednio lub pośrednio do okienka zalążka. Zazwyczaj, dopiero wówczas następuje wykształcenie gametofitów męskich. Dalsze udoskonalanie procesu zapłodnienia obejmowało redukcję plemników, które ze zdolnych do samodzielnego ruchu uwicionych komórek stały się niezdolnymi do ruchu pozbawionymi wici komórkami plemnikowymi. Komórki plemnikowe są biernie transportowane w pobliże komórki jajowej przez twór zwany łagiewką pyłkową. Współcześnie tę tendencję ewolucyjną prezentują rośliny nagonasienne i okrytonasienne.
Słownik
(gr. allos – inny) forma genu; jedna z wersji genu różniąca się od pozostałych sekwencją nukleotydów; zajmuje określone miejsce na chromosomie
inaczej samożywność; sposób odżywiania się, w którym organizm samodzielnie syntetyzuje złożone związki organiczne z prostych związków nieorganicznych przy udziale energii świetlnej lub energii chemicznej
część gametofitu, w której odbywa się powstawanie haploidalnych gamet służących do rozmnażania płciowego
pokolenie haploidalne rozmnażające się płciowo przez haploidalne gamety wytwarzane na drodze mitozy
gametofit, na którym występują tylko rodnie lub tylko plemnie
gametofit, na którym występują rodnie i plemnie
gametofit wytwarzający plemnie, w których powstają plemniki
gametofit wytwarzający rodnie, w których powstaje komórka jajowa
inaczej cudzożywność; sposób odżywiania się, w którym organizm pobiera związki organiczne ze środowiska zewnętrznego
gametangium żeńskie; część gametofitu wytwarzająca haploidalne gamety żeńskie – komórki jajowe
długa, nitkowata struktura wyrastająca z komórki wegetatywnej ziarna pyłku, przenosząca komórki plemnikowe w pobliże komórki jajowej
rodzaj zarodnika, z którego powstaje gametofit żeński
rodzaj zarodni wytwarzającej makrospory
rodzaj zarodnika, z którego powstaje gametofit męski
rodzaj zarodni, w której powstają mikrospory
organy odpowiedzialne za wzrost i rozwój rośliny; zaliczają się do nich: korzeń, łodyga i liście
gametangium męskie; część gametofitu, wytwarzająca haploidalne gamety męskie – plemniki
regularne i cykliczne następstwo jąder komórkowych o haploidalnej (1n, haploidalność) i diploidalnej (2n, diploidalność) liczbie chromosomów w cyklu życiowym organizmów rozmnażających się płciowo
gametangium żeńskie; cześć gametofitu, wytwarzająca haploidalną gametę żeńską – komórkę jajową
pokolenie diploidalne rozmnażające się bezpłciowo poprzez haploidalne zarodniki wytwarzane na drodze mejozy
komórki w woreczku zalążkowym u okrytonasiennych (zwykle dwie) z boku komórki jajowej; warunkują asymetrię otoczenia komórki jajowej; jedna z synergid po zapyleniu obumiera i do niej kieruje się łagiewka pyłkowa na swej drodze do powierzchni komórki jajowej
inaczej sporangium; część sporofitu, w której powstają haploidalne zarodniki służące do rozmnażania bezpłciowego