Przeczytaj
Genetyka
GenetykaGenetyka to jedna z najnowszych nauk w obrębie nauk biologicznych. Jest to równocześnie jedna z najprężniej rozwijających się dziedzin wiedzy, zajmująca się dziedzicznością i zmiennością organizmów. Termin „genetyka” wprowadził na początku XIX w. William Beatson. Ze względu na szeroki zakres badań prowadzonych w ramach tej nauki wyróżnia się kilka jej działów:
genetyka klasyczna – określana również jako mendlowska – zajmuje się badaniem mechanizmów dziedziczenia na podstawie widocznych cech organizmów (fenotypówfenotypów).
genetyka molekularna – dział genetyki opierający się na molekularnym mechanizmie dziedziczenia, budowie materiału genetycznego, jego sekwencji, zmienności i organizacji. Bada budowę oraz organizację genówgenów na poziomie struktury.
genetyka populacyjna – określa zmiany i zróżnicowanie genetyczne w obrębie danej populacji.
genetyka nowotworów – dział genetyki skupiający się na zaburzeniach regulacji cyklu komórkowego, prowadzących do zmian o charakterze nowotworowym.
Genetyka klasyczna (mendlowska)

Genetyka klasyczna opiera się w głównej mierze na badaniach Grzegorza Mendla. To właśnie jego odkrycia i koncepcje dały początek rozważaniom o prawach rządzących dziedziczeniem i przyczyniły się do dalszego rozwoju nauki. Kolejni naukowcy rozwijali myśl Mendla, wraz z nowymi odkryciami wprowadzając kolejne pojęcia i terminy. Nie bez powodu Grzegorza Mendla określa się więc jako twórcę genetyki.
Grzegorz Mendel od 1856 r. przez dziesięć lat przeprowadzał doświadczenia na mieszańcach roślin, przede wszystkim grochu zwyczajnego (Pisum sativum). Prowadząc badania, przestrzegał kilku zasad: używał wyselekcjonowanych czystych liniiczystych linii, ograniczał obserwację do nielicznych i wyraźnych cech, ustalał liczebność każdego fenotypu w kolejnych pokoleniach mieszańców oraz w potomstwie każdej rośliny.
Na podstawie swoich obserwacji Mendel opracował zasady dziedziczenia. Ich treść zmieniała się wraz z upływem czasu, w oparciu o nowe odkrycia. Współczesne ich wersje noszą nazwę praw Mendla:
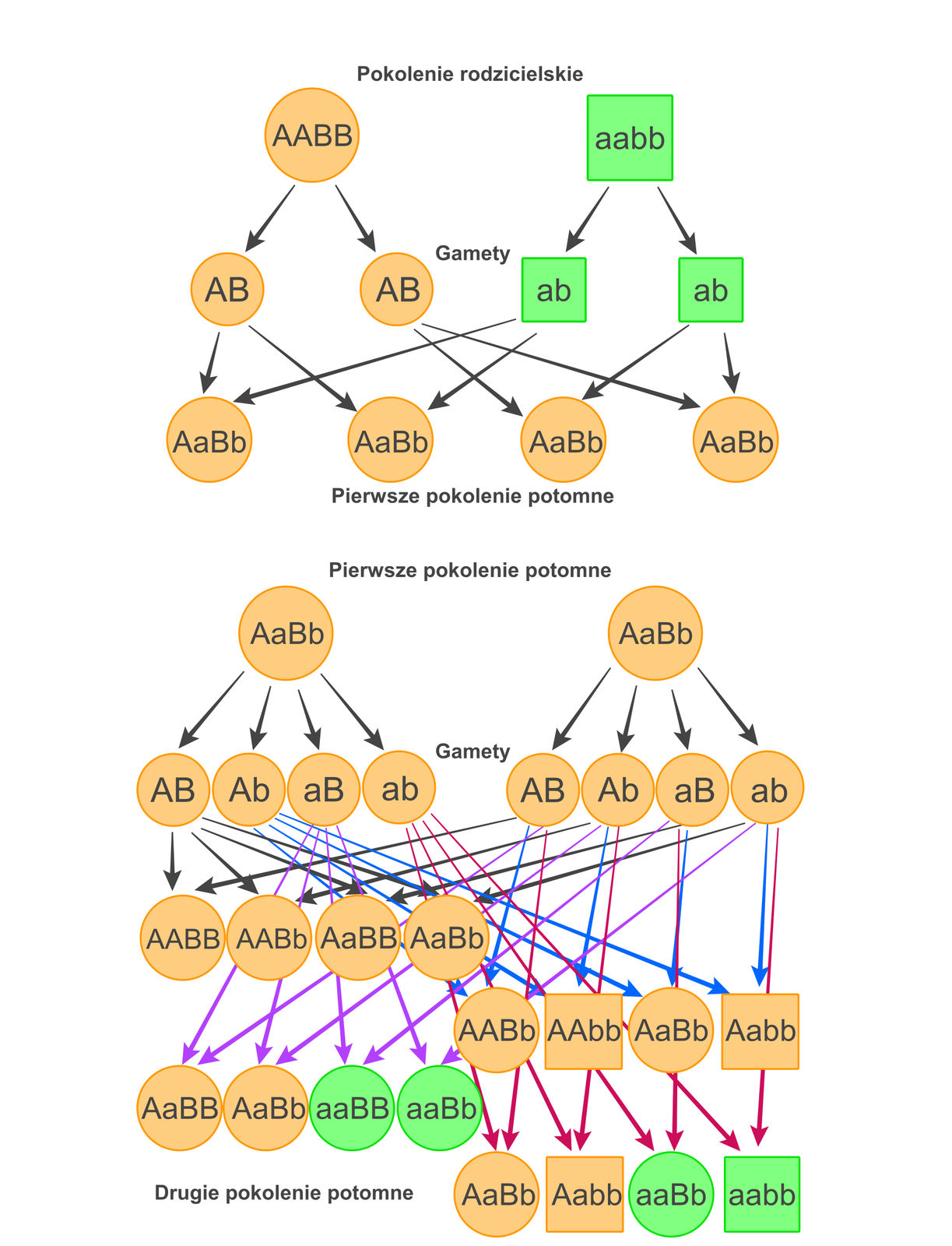
pierwsze prawo Mendla – prawo czystości gamet. Zgodnie z nim każda gameta, która wywarzana jest przez organizm, ma tylko jeden allelallel (wersję) z danej pary alleli genu.
drugie prawo Mendla – prawo niezależnej segregacji cech. Zgodnie z tym prawem cecha uwarunkowana przez daną parę alleli (gen) dziedziczona jest niezależnie od cechy warunkowanej przez inną parę alleli.
Podstawą genetyki mendlowskiej jest mejozamejoza, która zapewnia zmniejszenie o połowę ploidalności komórek – gamet, kompensując w ten sposób podwojenie informacji genetycznej występującej w momencie zapłodnienia. Mejoza stanowi wyjaśnienie prawa czystości gamet.

– A – allel dominujący,
– a – allel recesywny.
– A, B – allele dominujące,
– a, b – allele recesywne.
Więcej informacji na temat mejozy w materiale: Przebieg i znaczenie mejozyPrzebieg i znaczenie mejozy.

W pracach Mendla nie występowało pojęcie „gen” ani „allel”. Posługiwał się on terminem „czynniki dziedziczności” (lub „jednostki dziedziczności”). Uważał, że czynniki dziedziczności mogą wchodzić w skład trzech różnych par: AA, Aa lub aa, gdzie A to czynnik dominujący, podczas gdy a to czynnik recesywny. Linie czyste, na których pracował Mendel, to osobniki mające wyłącznie dwa czynniki recesywne (aa) – kwiaty o barwie białej, bądź dwa czynniki dominujące (AA), charakteryzujące kwiaty czerwone. Takie osobniki określane są jako homozygoty. W pierwszym pokoleniu potomnym (F1), uzyskanym po skrzyżowaniu osobników zawierających dwa czynniki dominujące AA (czerwona barwa kwiatów) z osobnikami o dwóch czynnikach recesywnych aa (biała barwa kwiatów), występowały wyłącznie osobniki zawierające zarówno czynnik dominujący jak i recesywny Aa, które nie różniły się wyglądem od osobników rodzicielskich posiadających dwa czynniki dominujące – miały kwiaty czerwone. Na podstawie swoich doświadczeń Mendel wywnioskował, że czynnik warunkujący czerwoną barwę kwiatów jest czynnikiem dominującym. Obecnie jest on określany jako allel dominującyallel dominujący, a czynnik warunkujący biały kolor kwiatów to allel recesywnyallel recesywny, który nie ujawni się w obecności allelu dominującego. Po skrzyżowaniu osobników z pokolenia F1 otrzymane rośliny w ok. 75% miały kwiaty barwy czerwonej, pozostałe zaś (ok. 25%) – kwiaty białe.
Sformułowane przez Mendla w 1865 r. podstawowe zasady genetyki nie od razu wzbudziły zainteresowanie w świecie nauki. Dopiero w 1900 r. ponownie odkryli je i potwierdzili (niezależnie od siebie, badając inne gatunki roślin) C. Correns (który prowadził badania na dziwaczku peruwiańskim (Mirabilis jalapa)), E. Tschermak (który prowadził badania na groszku pachnącym (Lathyrus odoratus)) i H. de Vries (który prowadził badania na wiesiołku dwuletnim (Oenothera biennis)).
Więcej na temat tych badań znajdziesz w e‑materiale: Rewolucja mendlowskaRewolucja mendlowska.
W 1905 r. Reginald C. Punnett zaproponował graficzną formę przedstawienia mechanizmu dziedziczenia. Od jego nazwiska nosi ona nazwę szachownicy Punnetta.
AA – osobnik mający dwa dominujące allele danego genu, czyli homozygota dominująca;
aa – osobnik mający dwa recesywne allele danego genu, czyli homozygota recesywna;
Aa – heterozygota, czyli osobnik mający zarówno allel dominujący, jak i recesywny danego genu.
Więcej na temat szachownicy Punnetta w e‑meteriale: Szachownica PunnettaSzachownica Punnetta.
Pojęcie „gen” pojawiło się dopiero w 1908 r. Opracował je Wilhelm Johannsen, skróciwszy nazwę „pangen”, zaproponowaną wcześniej przez Hugo de Vriesa. Całkowity zbiór genów osobnika to „genotyp”. Termin ten również został zaproponowany przez Johannsena. Przedstawił on także propozycję rozróżnienia między genotypem a fenotypem, który stanowi widoczne i mierzalne odzwierciedlenie genotypu. Pojęcie „allel”, oznaczające jedną z wersji genu, wprowadzili William Bateson oraz Edith Rebecca Saunders.
Geny zlokalizowane są na chromosomachchromosomach, które stanowią formę organizacji materiału genetycznego. Termin „chromosom” został po raz pierwszy użyty przez Heinricha Wilhelma Waldeyera w 1888 r., natomiast pierwsze obserwacje chromosomów datowane są na 1848 r. Chromosomy dzieli się na:
chromosomy płci (allosomy) – warunkujące płeć; ich rolę odkryła Nettie Stevens w 1905 r.;
autosomy – wszystkie chromosomy organizmu z wyjątkiem allosomów.
Zestaw wszystkich chromosomów w komórce to kariotyp. Allele danego genu zlokalizowane są w określonym miejscu (locuslocus) na chromosomie. Chromosomy, zawierające podobną informację genetyczną, a co za tym idzie, takie same lub różne allele genów, to chromosomy homologicznechromosomy homologiczne. W komórkach diploidalnych występują one parami. Chromosomy te rozdzielane są w procesie podziału komórki tak, że tylko jeden chromosom z pary trafia do komórki potomnej.
Fenotyp warunkowany jest przez genotyp. Próbę wytłumaczenia wpływu genów na wygląd organizmu próbowano tłumaczyć hipotezą jeden gen – jeden enzym. Według tej teorii gen koduje dany enzym, który z kolei wpływa na kształtowanie określonej cechy. Doświadczenie wykazujące słuszność tej tezy wykonali George Beadle i Edward Tatum na grzybie Neurospora crassa. Szybko pojawiły się jednak nowe informacje, które sprawiły, że hipotezę tę uznano za zbyt duże uproszczenie.
Więcej na ten temat w e‑materiale: Czym jest gen?
Chromosomowa teoria dziedziczenia

Chromosomową teorię dziedziczenia po raz pierwszy zaproponował w 1884 r. A. Weismann, sformułowali ją dokładnie na początku XX w. W. S. Sutton oraz T. Boveri, a doświadczalnie ugruntowali ją T. Morgan i jego współpracownicy. Wykazali oni, że chromosomy są zbiorami ułożonych liniowo genów, przy czym dany gen zajmuje zawsze jednakowe, ściśle określone miejsce (locus) na chromosomie. Ponadto geny jednego chromosomu są dziedziczone łącznie (są ze sobą sprzężone), natomiast geny znajdujące się w różnych chromosomach dziedziczą się niezależnie od siebie. Podczas koniugacji chromosomów homologicznych (wczesna profaza podziału I mejozy) może zachodzić zjawisko wzajemnej wymiany odpowiadających sobie odcinków chromatyd niesiostrzanych (crossing‑overcrossing‑over). Częstość zachodzenia crossing‑over jest wprost proporcjonalna do odległości między genami, tzn. im bliżej są one położone, tym silniej są sprzężonesprzężone, a prawdopodobieństwo rozdzielenia ich pomiędzy dwa różne chromosomy homologiczne jest mniejsze. Jednostką częstości zachodzenia crossing‑over jest centymorgancentymorgan (cM).
Chromosomowa teoria dziedziczenia przyczyniła się do poznania budowy chromosomów i biochemicznego zidentyfikowania nośnika informacji genetycznejinformacji genetycznej.
Więcej na ten temat w e‑materiale: Chromosomowa teoria dziedziczności MorganaChromosomowa teoria dziedziczności Morgana.
Odstępstwa od praw Mendla
Wraz z rozwojem genetyki oraz badaniem innych przykładów dziedziczenia okazało się, że istnieje szereg odstępstw od zaproponowanych przez Mendla zasad dziedziczenia. Zalicza się do nich:
Dominację niepełną – żaden z alleli genu kodującego określoną cechę nie dominuje w pełni nad drugim; heterozygota wykazuje fenotyp pośredni między fenotypem homozygoty dominującej i recesywnej. Więcej na ten temat w e‑materiale: Dominacja zupełna i częściowaDominacja zupełna i częściowa.
Allele wielokrotne – występują więcej niż dwa allele danego genu, chociaż diploidalna komórka danego organizmu zawiera jedynie dwa allele spośród kilku możliwych, a w gamecie występuje jeden allel. Więcej na ten temat w e‑materiale: Kodominacja i allele wielokrotneKodominacja i allele wielokrotne.
Zjawisko kodominacji – w fenotypie heterozygoty ujawniają się oba allele; nie można wyróżnić allelu dominującego i recesywnego, są one równorzędne. Więcej na ten temat doczytasz także w e‑materiale: Kodominacja i allele wielokrotneKodominacja i allele wielokrotne.
Plejotropowe działanie genów – dany gen wpływa na warunkowanie więcej niż jednej cechy fenotypowej. Więcej na ten temat w e‑materiale: PlejotropizmPlejotropizm.
Współdziałanie dwóch lub większej liczby genów – cecha fenotypowa warunkowana jest przez więcej niż jeden gen. Więcej na ten temat w e‑materiale: Współdziałanie genów – ćwiczeniaWspółdziałanie genów – ćwiczenia.
Dziedziczenie cech sprzężonych z płcią – geny (ich allele) odpowiedzialne za dane cechy znajdują się na chromosomach płci; najczęściej na chromosomie X. Ich wystąpienie zależy od układu chromosomów płci (XX, XY). Więcej na ten temat w e‑materiale: Cechy sprzężone z płciąCechy sprzężone z płcią.
Dziedziczenie pozajądrowe – warunkowane przez pozachromosomowy materiał genetyczny: mitochondrialny lub chloroplastowy (w przypadku roślin). Więcej na ten temat w e‑materiale: Dziedziczenie pozajądroweDziedziczenie pozajądrowe.
Zastosowanie genetyki klasycznej
Z czego wynika ogromne zainteresowanie świata nauki, zwłaszcza medycyny, genetyką? Wyniki badań genetycznych pozwalają stwierdzić np. dlaczego w jednej populacji częściej niż w innych występuje cukrzyca, jakie jest ryzyko rozwoju określonego nowotworu u danej osoby, a także które leki okażą się skuteczne w przypadku konkretnego chorego. Rozwój badań genetycznych łączy się więc i z medycyną populacyjną, i medycyną spersonalizowaną, nastawioną na dobór terapii ściśle dostosowanej do przypadku (w tym genotypu) danego chorego. Z wykorzystaniem inżynierii genetycznej powstają m.in. leki biologiczne, natomiast dzięki farmakogenetyce można określić typ leku i jego dawkę, które będą skuteczne i bezpieczne dla danego chorego.
Słownik
(gr. allos – inny) forma genu; jedna z wersji genu różniąca się od pozostałych sekwencją nukleotydów; zajmuje określone miejsce na chromosomie
oznaczany jest dużą literą; maskuje obecność allelu recesywnego w heterozygocie (allel recesywny nie może się ujawnić)
oznaczany jest małą literą (np. a); jest to jedna z wersji danego genu (np. A), która ujawnia się w homozygocie recesywnej (np. aa). W heterozygocie (np. Aa) cecha warunkowana przez ten allel nie ujawnia się, co oznacza, że allel recesywny jest maskowany przez allel dominujący
1 cM odpowiada 1% rekombinacji (częstości zachodzenia crossing‑over); termin cM wprowadzono na cześć T. H. Morgana, twórcy chromosomowej teorii dziedziczności, który opisał zjawisko sprzężenia genów
(gr. chrṓma – barwa, sṓma – ciało); struktury zawierające materiał genetyczny komórki
jeden z nich pochodzi od ojca, a drugi od matki; chromosomy te mają tę samą długość ramion, takie samo położenie centromeru, a znajdujące się w tym samym locus geny (allele) warunkują tę samą cechę, np. barwę oczu; podczas mejozy chromosomy homologiczne łączą się, tworząc biwalenty
proces polegający na wzajemnej wymianie odpowiadających sobie odcinków chromatyd chromosomów homologicznych; rezultatem crossing‑over jest przetasowanie genów pochodzenia matczynego i ojcowskiego, prowadzące do powstania nowych, przypadkowych kombinacji alleli w ramach jednego chromosomu; crossing‑over zachodzi w profazie I mejozy
(gr. phaínomai – pojawiam się, týpos – odbicie); zespół anatomicznych, fizjologicznych i biochemicznych cech organizmu o podłożu dziedzicznym, które można obserwować i mierzyć
podstawowa jednostka dziedziczności; odcinek , w obrębie którego jest zapisana informacja o budowie białka lub RNA
(gr. genétēs, génesis – 'rodzic', 'zrodzony'); nauka zajmująca się badaniem dziedziczności i zmienności organizmów
informacja zawarta w sekwencji kwasów nukleinowych ( lub w przypadku niektórych wirusów RNA), która jest przekazywana organizmom potomnym
zespół osobników homozygotycznych powstały w wyniku chowu wsobnego
l. poj.: locus, l. mn.: loci; określone miejsce na chromosomie, które zajmuje gen
(gr. meiosis – zmniejszenie) podział redukcyjny jądra komórkowego, prowadzący do wytworzenia haploidalnych komórek płciowych, czyli gamet, u zwierząt tkankowych (oogeneza, spermatogeneza), zarodników u mszaków i paprotników oraz ich odpowiedników u roślin nasiennych (sporogeneza); u organizmów niższych (np. glonów, pierwotniaków) mejoza nie musi być związana z produkcją gamet bądź zarodników i zachodzi w różnych fazach cyklu życiowego (koniugacja, przemiana pokoleń)
podstawowe reguły dotyczące przekazywania cech warunkowanych genetycznie podczas rozmnażania płciowego, sformułowane w 1865 r. przez G. J. Mendla w wyniku wieloletnich badań nad dziedziczeniem u samopylnej rośliny – grochu zwyczajnego
łączne dziedziczenie się dwóch lub więcej genów (i warunkowanych przez nie cech)